|
В 1965 году молодой британский биолог-эволюционист Уильям Гамильтон предложил научному миру свое видение того, почему в животном мире есть альтруисты, жертвующие своими интересами ради других, и агрессоры, которых вообще не интересуют проблемы и заботы окружающих (позднее теорию Гамильтона популяризовал Ричард Докинз в своей книге «Эгоистичный ген»). Несмотря на почти полувековое существование, эта теория во многом так и осталась теорией, слабо подкрепленной реальными примерами из жизни. Ученые из Оксфордского университета доктор Стюарт Вест (Stuart A. West) и доктор Энди Гарднер (Andy Gardner) на страницах журнала Science приводят обзор самых последних исследований того, как работает теория Гамильтона.
Предисловие
Сбор Ли Алан Дугаткин провозгласил Гамильтона Эйнштейном в биологии, потому что K =½ r, это «своего рода e = mc2 для эволюционной биологии». Правда, в Европе и Америке так думают не все. Когда в 2010 году Новак (M.A. Nowak), Тарнита (C.E. Tarnita)и О. Уилсон (E.O. Wilson) объявили короля голым, одни назвали эту статью шагом вперед, другие – безумием. В России только единицы с самого начала отнеслись к этой теории со святым гневом и непримиримой критикой. Я, например, могу назвать только двоих. Это этолог Евгений Николаевич Панов и энтомолог Владилен Евгеньевич Кипятков. Глядя на многих других, я уже устал удивляться, как легко первый встречный жулик делается властителем дум бесхитростного и доверчивого русского человека. Как склонен даже ученый русский человек верить всему, даже не пощупав. Вместо того, чтобы броситься в поле или в лабораторию и проверить чужой результат, он становится примитивным переписчиком чужих книжек и статей. Ничего принципиально не изменилось и после статьи авторитетного О. Уилсона с коллегами. На сайте Derkeiler.com Гамильтона перевели из Эйнштейнов в Ньютоны, в том смысле, что Гамильтон не ошибся, и его закон является частным случаем дарвиновской теории. Такое толкование места Гамильтона в истории науки напрашивается из вывода, который делают M.A. Nowak, C.E. Tarnita и E.O. Wilson. «Будучи лишь одной из возможных моделей эволюции, - говорят они, - теория итоговой приспособленности оказалась ошибочно признана справедливой для всей сферы эволюции социального поведения». В этом небольшом эссе я хочу показать, что теория Гамильтона не соответствует генетической реальности, что особенно удивительно для генетической теории, и что Гамильтон не имеет ясного представления о том, что пытается объяснить.
История вопроса
Сбор Теория альтруизма Уильяма Гамильтона появилась на свет, чтобы объяснить недоразумение, обнаруженное Чарльзом Дарвином. Во-первых, Дарвин рассматривал благородство, которое принято считать синонимом альтруизма, не как общечеловеческое свойство, а как наследственное свойство избранных индивидов. Во-вторых, он традиционно рассматривал альтруизм, как разновидность самоубийства. И вот, опираясь на эти представления он делает вывод, что благородный индивид «часто вовсе не оставлял потомства, способного унаследовать его благородную натуру», потому что «наиболее мужественные, в среднем должны погибать в большем количестве, нежели менее храбрые». Проблема состояла в том, что естественный отбор по разумению Дарвина должен был уничтожить альтруизм, но этого по какой-то причине не произошло. И сначала Холдейн, а потом Гамильтон предложили очень простое и, на первый взгляд, даже гениальное объяснение этого парадокса.(из статьи В. Унраву,2012. (http://paseka-ru.narod.ru/sbornik-gen-unrau.html).
Совокупная приспособленность альтруиста.
Совокупная приспособленность альтруиста; ai – прямой вклад или индивидуальная приспособленность альтруиста; x – приспособленность родственников альтруиста. Вообще говоря, это была не новая идея. То, что наследственный материал, выраженный хорошим признаком, есть не только у особо одаренного индивида, но и у его родственников, было хорошо известно и Дарвину.
«Опыт сельских хозяев, - пишет он, - показал, что, сохраняя потомство от стада, в котором было найдено ценное животное, можно достичь накопления желаемых признаков, даже в том случае, если избранное животное было убито, не оставив потомства».
Так вот, Гамильтон зацепился за эту идею, и предложил считать приспособленность альтруистического организма не по числу его собственных потомков, как принято считать в популяционно-генетической теории отбора, а по общему количеству аллелей альтруизма, которые останутся после него в популяции. Такая приспособленность получила название совокупной (inclusive fitness)
.
i = ai + x,
Правда, аллель альтруизма есть не у всех родственников. Поэтому их вклад в совокупную приспособленность альтруиста зависит от частоты аллеля альтруизма, с которой он у них встречается.
Гамильтон говорит, что отбор благоприятствует генам альтруистического поведения, «если выгода в репродуктивных шансах у получателя помощи, поделенная на потери репродуктивных шансов у «помощника», составляет величину, большую, чем обратное значение степени родства особей».
Это правило выражается неравенством R > c/b, где R – степень родства, с (cost) – затраты альтруиста, b (benefit) – выигрыш получателя альтруистической помощи.
Собсвенно говоря, Гамильтон в 1964 году перевел идею Холдейна за 1955 год на математический язык, и в этом состоит заслуга, которую ему приписывают.
Степень родства.
Степень или коэффициент родства Гамильтон рассчитывает по упрощенной формуле Райта
RXY=∑0,5n.
R в этой формуле – коэффициент генетического сходства между особями; n – генерационное расстояние между особями; 0,5 – доля аллеля в генотипе.
Рассчитывая по этой формуле степень родства, получают, что у братьев 0,5 общих аллелей, у племянников с альтруистом – 0,25, у двоюродных братьев – 0,125 и т.д.
Правда, упрощенная формула Райта не учитывает инбридинга и не считает одинаковые аллели у неродственников тождественными, поэтому, строго говоря, для теории Гамильтона коэффициент родства не равен частоте аллеля:
R q или p.
Здесь q и p – частоты разных аллелей (А и а) одного гена.
Ведь для того, чтобы в популяции появился альтруистический генотип (аа), необходим кровнородственный брак между генотипами Аа и Аа или гетерозиготы Аа должны быть достаточно широко распространены в популяции. И при широком распространении Аа в популяции частота аллеля а среди родственников альтруиста будет гораздо выше, чем считает Гамильтон.
Тем не менее, это ничего не меняет в принципе.
Практическим следствием этой модели, по мнению Гамильтона, является то, что для сохранения аллеля альтруизма в популяции, альтруист должен спасти некоторое количество своих родственников, чтобы среди них случайно оказался носитель аллеля альтруизма.
Узловая идея Гамильтона и Холдейна вытекает из того факта, что у родственников альтруиста тоже должны быть аллели альтруизма, которые будут продолжать род альтруистов. .(из статьи В. Унраву,2012. (http://paseka-ru.narod.ru/sbornik-gen-unrau.html).
Альтруизм в семьях пчел, с подходом к теории Гамельтона.
Например, известно, что в семье медоносных пчел пчелинная матка создает многочисленные копии рабочих пчел (роботов),из которых отдельные копии при обнаружении опасности для жизни семьи, в защиту своего гнезда и спасая своих сестер жертвуют своей жизнью.. Родственники по крови,рабочии пчелы по определению, очень похожи друг на друга. Так что, хотя маленькие рабочие пчелы и жертвовали собственной жизнью, своими действиями они потенциально спасали сотни родственников по крови. Современным языком мы бы сказали, что рабочие пчелы помогали своим кровным родственникам, потому что между кровными родственниками существует генетическая связь. Помогая своим кровным родственникам, вы косвенно способствуете воспроизведению копий собственных генов – генов, которые принадлежат вашей семье.
(Из сайта paseka-ru, 2012, Ю. Третьяков.)
Альтруизм или? - злобность копий отдельных групп пчел.
В литературных источниках по генетике пчел нет понятия гена альтруизма, а принят как ген злобности (агрессии), т.е к любым нарушителям в попытке вмешательство в гнездо семьи пчел. Однако, у пчеловодов практиков могут возникнуть противоречие о злобности пчел разных пород. Например, пчелы Среднерусские местные и метисы или Башкирские очень злобные и агрессивны. Они нападают с целью ужалить не только вблизи гнезда, но и с расстояния 5-10 метров и более (50 м.) от улья и даже в близи пасеки. Для защиты от таких злых пчел пчеловоды применяют плотную одежду, сетку для лица, перчатки для защиты рук и обувь такую, высокие батинки, керзовые или резиновые сапоги.(рис.).

В тоже время такие пчелы, чистых пород первого поколения, как Карпатские, Карника, Басфат относятся к не злобным. С этими породами в теплую солнечную погоду и наличия взятка с медоносов пчеловоды часто могут проводить работы в открытых ульях не боясь переставлять или вынимать сотовые рамки с пчелами и стряхивать пчел с рамок и т.п. Они иногда осматривают пчел без защитных сеток и костюмов в одних шортах. Следует отметить, что на пасеке не должны находиться агрессивные породы и помеси агресивных с не агрессивными.
Какие особи рабочих пчел в семье могут быть агрессивными.
Родоначальницей в каждой семье пчёл является плодная матка, которая живет несколько лет только в составе семьи и ежегодно в период цветения медоносов. с весны до осени, откладывает в ячейки сот своего гнезда множество яиц оплодотворенных сперматозоидами трутней. Из яиц черз 3 дня образуются личинки из которых рабочие пчёлы воспитывают многотысячное потомство женских рабочих особей-сборщиц кормовых запасов из нектара и пыльцы.
В активный период жизнедеятельности продолжительность жизни одного поколения рабочих особей составляет в среднем около 30 -35 дней. Одно поколение образуется рт числа яиц отложенных маткой пчел за сутки. Число поколений зависит от яйценоскости матки и факторов внешней среды и медоносов.. Наибольшее число рабочих особей пчелы выращивают к сбору зимних запаов меда. К этому периоду численость семьи может составлять от 30 до 60 тысяч и более особей. Из этого числа охрана гнезда из стороэевых пчел может составлять около 0,5- 1%. При защите гнезда могут принять участие 10-20% особей. В Защитуе пчелы используют выдвижное жало на конце брюшка.
Жало имеется только у матки и рабочих пчел. Яд у них выделяется железой с 6-7 дневного возраста, но наиболее активно на 10-18 день. Накопление яда наблюдается с 7 до 20 дневного возраста. В железе накапливается около 0,2 мг яда. Следовательно, начиная с 10-18 дней жизни каждая взрослая рабочая особь может включается к охране своего семейного гнезда. В этом возрасте большинство особей становятся злыми и агрессивными при нападении на нарушителя жизнедеятельности семьи в улье. Для охраны гнезда подключаются особи на 23 день своей жизни и в течении 7 дней находится на страже у летка при входе в улей. Сторожевые пчелы предупреждают других членов семьи об опасности и вызывают подмогу. Подмога из многочисленных агрессивных особеи нападают не щадя своей жизни ради спасения своих генетических сестер, матки, расплода и кормов в своем гнезде. Такая особенность признака агреси и самопожертвования в защиту членов семьи и гнезда заложена у каждой рабочей особи в аллели гена хромосомы.
Число хромосом в клетках тела и оловых органах пчёл
|
Связь между родителями и потомством проявляется в информационной наследственности, которая передается от одного поколения другому через гены расположенные в определенной последовательности в хромосомах каждой особи. В одной хромосоме каждый ген занимает определенное место и контролирует один признак или несколько признаков наследственности. Известно, что все живые существа состоят из клеток, а в ядре клетки находятся хромотиновые зёрна, из которых путем деления образуются хромосомы. В каждой клетке любого организма у различных видов животных содержится, как правило, определенное число хромосом. По числу хромосом определяется пол животного или насекомого. У медоносной пчелы пол определяется с развитием рабочих пчёл и маток из оплодотворенных яиц, а трутней из неоплодотворенных яиц. Эту закономерность впервые открыл великий силезский пчловод Ян Дзержон в 1845 году (Шаскольский, 1968).
Клетки организма (соматические) женских особей пчёл имеют диплоидный набор хромосом (2x16=32), то есть двойной. Соматические клетки мужских особей (трутней) содержат гаплоидный набор, т.е. они происходят из гамет матки и в своих клетках тела имеют по 16 хромосом (n - один геном). В половых же органах матки и пчелы-трутовки развитые яйца в яичниках содержат по 16 хромосом. В семяприемнике плодной матки и в половых органах половозрелого трутня, сперматозоиды содержат по 16 хромосом (рис. 6).
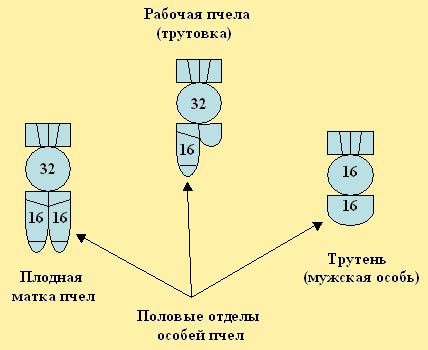
Рис. 6. Число хромосом в клетках пчёл
В результате из оплодотворенных яиц развиваются диплоидные женские особи-матки и рабочие пчёлы, а из неоплодотворенных яйцеклеток (партеногенез) развиваются мужские особи трутни (рис. 7), имеющие гаплоидный (одинарный) набор хромосом.
Иногда, из неоплодотворенного яйца может развиться диплоидная особь с 32 хромосомами. Такие случаи можно наблюдать у осиротевшей длительное время семьи пчел. Из яиц пчёл трутовок, пчёлы выводят матку неспособную к оплодотворению трутнями.
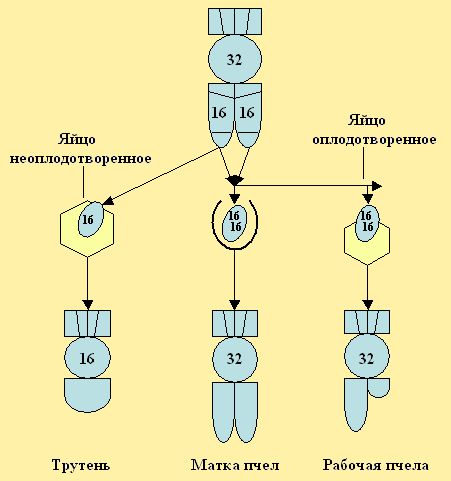
Рис. 7. При откладке, плодной маткой , в ячейки сот, оплодотворенных яиц, развиваются рабочие пчёлы и матки, их тело имеет 32 хромосомы, трутень - из неоплодотворенного яйца - 16.
В хромосомах распложены гены, которые содержат наследственную информацию. Следовательно, ген является единицей наследственной информации. Ген определяет развитие у пчёл одного или нескольких признаков. Число генов во много раз больше, чем хромосом. Хромосома передается по наследству как единое целое при коньюгации (процесс - сближения) и расхождение в мейозе, сохраняя свою целостность, а потому все гены, локализованные в одной хромосоме, обычно наследуются совместно. У пчел, каждая хромосома содержит один из аллелей гена.
Гены взаимоисключающих признаков составляют пару аллелей. Например, признак желтой окраски и темной (серой) окраски или признаки злобности и миролюбия у рабочих пчел, такие признаки в мейозе оказываются в разных гаметах.
Разные аллельные формы возникают при мутации (изменение наследственного материала) гена, свойственного дикому (нормальному) типу, где в паре аллелей один ген нормальный, т. е. неизменный, а другой измененный, то есть мутантный. Мутации, одного аллеля, нормального гена могут возникать несколько раз, в результате образуется серия аллелей. Это явление называют множественным аллелизмом - т. е. более двух аллельных форм гена. Ряд мутаций могут находиться в разных доминантно-рецессивных отношениях друг к другу и могут вызывать появления серии аллелей (C1, C2, C3,...Cn, которые составляют ряд.
Мутации по значению для организма пчёл могут быть полезными, вредными, нетральными, а по характеру возникновения - естественные или при искусственном оплодотворении.
Для обозначения аллелей в генетике принята буквенная символика, в которой доминантный признак (признак проявляется у гибридных особей подавляя внешней рецессивный) обозначают заглавной буквой А, В или С и т.д. Рецессивные признаки (внешне скрытые) обозначаются строчными буквами - а, в, с и т. д. Если, доминантный признак не полностью подавляет рецессивный, то такое явление называют неполным доминированием.
У медоносных пчёл пол определяет не вся хромосома, а только её участок
- ген, он является элементарной единицей наследственной информации.
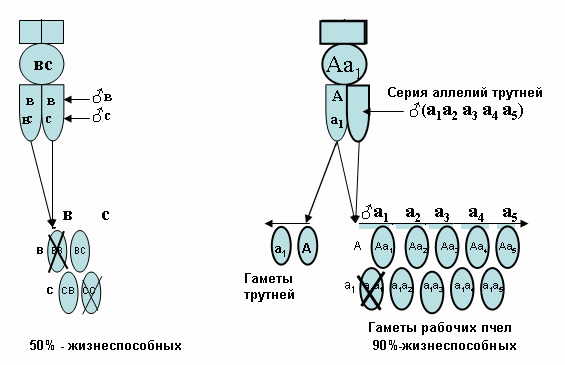
О. Маккензен и Робертс с 1951 года (Шаскольский, 1966) обозначили ген пола у пчелы, как и у наездника хабробракона (из семейства наездников браконид), через Х, а его аллели через Ха, Хb, Xc...... Для простоты аллели принято обозначать, как а, в, с, и т.д., помня, что это аллели гена Х (Шаскольский, 1966).
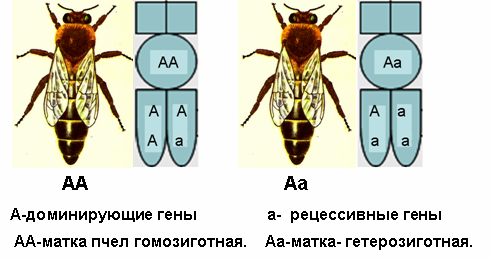
Известно, что каждый ген имеет множество различных типов: а, а2, а3,...а6......а12, которые называют половыми аллелями. Если в оплодотворенном яйце встречаются 2 аллеля, например, как а2а8 или а5а11 всегда разовьется самка, а если - а2а2 или а8а8 - разовьется самец, но он будет для пчёл нежизнеспособным, как правило пчёлы такие яйца поедают. Если рабочая пчела или матка имеют два одинаковых аллеля, например, АА или аа т.е. гомологичную пару, то они будут называться гомозиготными по паре одинаковых аллелей, а если аллели различны, как Аа, Вв, Сс и т.д. то такие особи называют гетерозиготными.
Общественная жизнь современных медоносных пчёл выражается во взаимном сотрудничестве - передаче друг другу информации, корма, постоянного контакта с маткой, коллективного выполнения всех работ обеспечивающих жизнь свою и всех последующих поколений.
Современная система наследование пола использует ген с обозначением «X» по О. Маккензен (рис.12), либо с обозначением «S» (рис.13). S от латинского слова sexus - пол по Шаскольскому (1990). Эти обозначения ген могут иметь целую серию множественных аллелей (Sa, Se, Sg, Si ......). Матка всегда гетерозиготна и может иметь гены по её матери и отцу, а после спаривания с трутнями в семяприемнике может иметь целую серию одинаковых или разных аллелей.
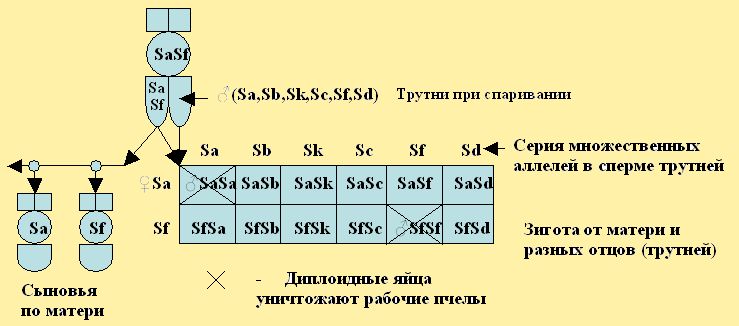
Рис. 13. Модель наследования гена пола от плодной матки, дающая часть гомозиготных яиц из которых могли бы развиться диплоидные трутни.
6 .1. Законы наследования и изменчивости признаков
медоносых пчёл.
6.1.3. Цитологические основы закономерностей наследования
Связь между поколениями особей пчёл при половом размножении осуществляется через половые клетки (гаметы), которые несут материальные наследственные факторы - гены. Гены определяют развитие того или иного признака они расположены в хромосомах. В клетках женских особей пчел, как уже отмечалось ранее, число хромосом составляют 32 (две пары гамет), а у мужских (трутней) 16 (одна пара гамет). Половые клетки женских и мужских особей составляют одну пару хромосом. Две пары хромосом возможны при слиянии женской яйцеклетки и мужской клетки (сперматозоида) в зиготе. Зигота может произойти от слияния женской яйцеклетки и сперматозоида, содержащие одинаковые гены и разные. Например, когда яйцеклетка с геном – А соединится сперматозоидом имеющим ген – А, зигота будет АА, а если яйцеклетка А соединится со сперматозоидом – а то зигота будет Аа. Гомозиготная пчелиная матка АА или аа имеет яйца из гамет А или а, и трутни народившиеся от таких маток будут иметь гаметы А или а.
При рассмотрении цитологических основ моногибридного скрещивания в первом поколении F1 мы наблюдаем в зиготе соединение гамет и расхождение их в мейозе. Это специфическое деление клеток, при котором образуются половые клетки для второго поколения F2 (рис. 25.1). На представленном рисунке 25.2. на моделях пчёл и символах, можно рассмотреть, что у родителей первого поколения F1  АА – желтой окраски и трутня АА – желтой окраски и трутня  а – черной окраски все потомство будет единообразно, т.е. – желтое. Во втором поколении F2 произошло расщепление в желтых особях на желтых и черных в соотношении 3:1, где три будут нести желтую окраску, а одна особь – черную. а – черной окраски все потомство будет единообразно, т.е. – желтое. Во втором поколении F2 произошло расщепление в желтых особях на желтых и черных в соотношении 3:1, где три будут нести желтую окраску, а одна особь – черную.
В генетике гетерозиготная особь носит название гибрид. Потомка, полученного от скрещивания родителей разных пород, называют метисом.
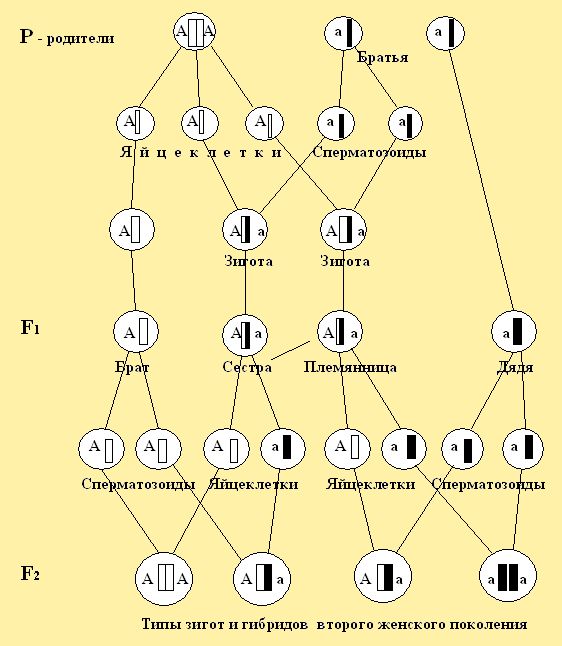
Рис. 25.1. Цитологические основы моногибридного расщепления, где расхождение пар гамет в мейозе и объединение их при оплодотворении в типах зигот потомства пчел.
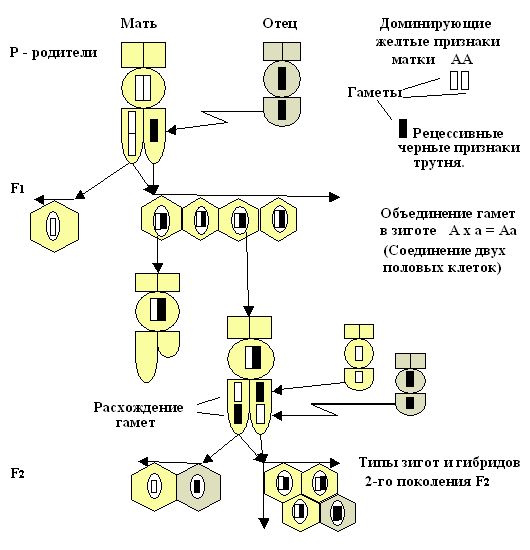
Рис. 25.2. Цитологические основы моногибридного расщепления гамет в потомстве от гомозиготной матки желтого доминирующего признака и гомозиготного трутня черного рецессивного признака (на моделях пчел). Расхождение пар гамет в яйцеклетках при мейозе и объединение их при оплодотворении в типах зигот во втором поколении гибридов.
|
Сайт создан в системе
uCoz
|
|
|