Статьи раздела Генетика
Статья Виктора Унрау
Один ген – один признак
Записки о теории Гамильтона и об альтруизме 2
СCодержание
Предисловие
История вопроса
Совокупная приспособленность
Примеры применения
Один ген – один признак
Совокупная приспособленность и популяционная генетика
Эусоциальность и альтруизм
Нужна ли гипотеза Гамильтона?
Что такое риск?
Для чего нужен риск?
Моббинг
Альтруисты Дарвина
Есть ли ген альтруизма?
Анализ результатов Эбштейна
Что такое альтруизм?
Двоякая природа альтруизма
Выводы
Вводная часть о теории Гамельтона
Термин альтруизм введён в противоположность эгоизму
В 1965 году молодой британский биолог-эволюционист Уильям Гамильтон предложил научному миру свое видение того, почему в животном мире есть альтруисты, жертвующие своими интересами ради других, и агрессоры, которых вообще не интересуют проблемы и заботы окружающих (позднее теорию Гамильтона популяризовал Ричард Докинз в своей книге «Эгоистичный ген»). Несмотря на почти полувековое существование, эта теория во многом так и осталась теорией, слабо подкрепленной реальными примерами из жизни. Ученые из Оксфордского университета доктор Стюарт Вест (Stuart A. West) и доктор Энди Гарднер (Andy Gardner) на страницах журнала Science приводят обзор самых последних исследований того, как работает теория Гамильтона.
(Из стаьей о теории Гамельтона).
Например, в семье медоносных пчел пчелинная матка создает многочисленные копии рабочих пчел (роботов),из
которых отдельные копии при обнаружении опасности для жизни семьи, в защиту своего гнезда и спасая своих сестер жертвуют своей жизнью.. Родственники по крови,рабочии пчелы по определению, очень похожи друг на друга. Так что, хотя маленькие рабочие пчелы и жертвовали собственной жизнью, своими действиями они потенциально спасали сотни родственников по крови. Современным языком мы бы сказали, что рабочие пчелы помогали своим кровным родственникам, потому что между кровными родственниками существует генетическая связь. Помогая своим кровным родственникам, вы косвенно способствуете воспроизведению копий собственных генов – генов, которые принадлежат вашей семье. Автор сайта paseka-ru.Ю. Третьяков
Предисловие
Ли Алан Дугаткин провозгласил Гамильтона Эйнштейном в биологии, потому что K =½ r, это «своего рода e = mc2 для эволюционной биологии».
Правда, в Европе и Америке так думают не все. Когда в 2010 году Новак (M.A. Nowak), Тарнита (C.E. Tarnita)и О. Уилсон (E.O. Wilson) объявили короля голым, одни назвали эту статью шагом вперед, другие – безумием.
В России только единицы с самого начала отнеслись к этой теории со святым гневом и непримиримой критикой. Я, например, могу назвать только двоих. Это этолог Евгений Николаевич Панов и энтомолог Владилен Евгеньевич Кипятков.
Глядя на многих других, я уже устал удивляться, как легко первый встречный жулик делается властителем дум бесхитростного и доверчивого русского человека. Как склонен даже ученый русский человек верить всему, даже не пощупав.
Вместо того, чтобы броситься в поле или в лабораторию и проверить чужой результат, он становится примитивным переписчиком чужих книжек и статей.
Ничего принципиально не изменилось и после статьи авторитетного О. Уилсона с коллегами.
На сайте Derkeiler.com Гамильтона перевели из Эйнштейнов в Ньютоны, в том смысле, что Гамильтон не ошибся, и его закон является частным случаем дарвиновской теории.
Такое толкование места Гамильтона в истории науки напрашивается из вывода, который делают M.A. Nowak, C.E. Tarnita и E.O. Wilson.
«Будучи лишь одной из возможных моделей эволюции, - говорят они, - теория итоговой приспособленности оказалась ошибочно признана справедливой для всей сферы эволюции социального поведения».
В этом небольшом эссе я хочу показать, что теория Гамильтона не соответствует генетической реальности, что особенно удивительно для генетической теории, и что Гамильтон не имеет ясного представления о том, что пытается объяснить.
История вопроса
Теория альтруизма Уильяма Гамильтона появилась на свет, чтобы объяснить недоразумение, обнаруженное Чарльзом Дарвином.
Во-первых, Дарвин рассматривал благородство, которое принято считать синонимом альтруизма, не как общечеловеческое свойство, а как наследственное свойство избранных индивидов. Во-вторых, он традиционно рассматривал альтруизм, как разновидность самоубийства.
И вот, опираясь на эти представления он делает вывод, что благородный индивид «часто вовсе не оставлял потомства, способного унаследовать его благородную натуру», потому что «наиболее мужественные, в среднем должны погибать в большем количестве, нежели менее храбрые».
Проблема состояла в том, что естественный отбор по разумению Дарвина должен был уничтожить альтруизм, но этого по какой-то причине не произошло.
И сначала Холдейн, а потом Гамильтон предложили очень простое и, на первый взгляд, даже гениальное объяснение этого парадокса.
ССовокупная приспособленность
Узловая идея Гамильтона и Холдейна вытекает из того факта, что у родственников альтруиста тоже должны быть аллели альтруизма, которые будут продолжать род альтруистов.
Вообще говоря, это была не новая идея. То, что наследственный материал, выраженный хорошим признаком, есть не только у особо одаренного индивида, но и у его родственников, было хорошо известно и Дарвину.
«Опыт сельских хозяев, - пишет он, - показал, что, сохраняя потомство от стада, в котором было найдено ценное животное, можно достичь накопления желаемых признаков, даже в том случае, если избранное животное было убито, не оставив потомства».
Так вот, Гамильтон зацепился за эту идею, и предложил считать приспособленность альтруистического организма не по числу его собственных потомков, как принято считать в популяционно-генетической теории отбора, а по общему количеству аллелей альтруизма, которые останутся после него в популяции. Такая приспособленность получила название совокупной (inclusive fitness)
.
i = ai + x,
где i – совокупная приспособленность альтруиста; ai – прямой вклад или индивидуальная приспособленность альтруиста; x – приспособленность родственников альтруиста.
Правда, аллель альтруизма есть не у всех родственников. Поэтому их вклад в совокупную приспособленность альтруиста зависит от частоты аллеля альтруизма, с которой он у них встречается.
Гамильтон говорит, что отбор благоприятствует генам альтруистического поведения, «если выгода в репродуктивных шансах у получателя помощи, поделенная на потери репродуктивных шансов у «помощника», составляет величину, большую, чем обратное значение степени родства особей».
Это правило выражается неравенством R > c/b, где R – степень родства, с (cost) – затраты альтруиста, b (benefit) – выигрыш получателя альтруистической помощи.
Собственно говоря, Гамильтон в 1964 году перевел идею Холдейна за 1955 год на математический язык, и в этом состоит заслуга, которую ему приписывают.
Степень или коэффициент родства Гамильтон рассчитывает по упрощенной формуле Райта
RXY=∑0,5n.
R в этой формуле – коэффициент генетического сходства между особями; n – генерационное расстояние между особями; 0,5 – доля аллеля в генотипе.
Рассчитывая по этой формуле степень родства, получают, что у братьев 0,5 общих аллелей, у племянников с альтруистом – 0,25, у двоюродных братьев – 0,125 и т.д.
Правда, упрощенная формула Райта не учитывает инбридинга и не считает одинаковые аллели у неродственников тождественными, поэтому, строго говоря, для теории Гамильтона коэффициент родства не равен частоте аллеля:
R q или p.
Здесь q и p – частоты разных аллелей (А и а) одного гена.
Ведь для того, чтобы в популяции появился альтруистический генотип (аа), необходим кровнородственный брак между генотипами Аа и Аа или гетерозиготы Аа должны быть достаточно широко распространены в популяции. И при широком распространении Аа в популяции частота аллеля а среди родственников альтруиста будет гораздо выше, чем считает Гамильтон.
Тем не менее, это ничего не меняет в принципе.
Практическим следствием этой модели, по мнению Гамильтона, является то, что для сохранения аллеля альтруизма в популяции, альтруист должен спасти некоторое количество своих родственников, чтобы среди них случайно оказался носитель аллеля альтруизма.
Вот так выглядит гипотеза Гамильтона.
ПрПримеры применения
ВВС сообщает: ученые «обнаружили, что монархи-убийцы интуитивно соблюдали «правило Гамильтона».
Жертвами королевского террора чаще всего были двоюродные братья и сестры. Генрихи IV и VIII убили по пять кузенов и кузин. Елизавета I, казнила двух кузенов и одну кузину – Марию Стюарт.
Почему английские монархи казнили преимущественно кузенов?
«Биологи говорят, - рассказывает ВВС, - что, если, например, монарх убивает двух родных братьев, то генетическая линия прерывается, поскольку каждый из убитых братьев будет обладать половиной генов убийцы. Если же убить пять кузенов, как это сделал Генрих VIII, то семья потеряет лишь треть своих генов, и династия сохранится».
Так английские короли от Эдварда III до Елизаветы I «сумели сохранить и трон, и свою генетическую линию» благодаря «интуитивному соблюдению правила Гамильтона».
Я бы легко поверил, что какой-то журналист сочинил эту чепуху, и для убедительности сослался на ученых из Кембриджа, если бы сами ученые из других научных центров не сочиняли такой же вздор.
Много такого рода фактов, которые якобы подтверждают, что Гамильтон был прав, можно найти, например, у Бэрэша.
Оказывается в племенах масаи, зулу и свази «отцы и сыновья вынуждены конкурировать меж собой» из-за жен по генетическим соображениям.
Дело в том, что за жену нужно платить калым «от 10 до 20 голов скота», и отец ломает голову: или взять еще одну жену, или женить сыновей, потому что всем хочется жениться, а стадо одно.
И думают все не о плотском, а о вечном!
Отцу выгоднее «производить детей, поскольку каждый из них нес бы в себе половину его генов», а внуки только ¼, «ведь родство разбавляется наполовину при каждом следующем поколении». А сыновья «предпочитают собственных детей (½) своим братьям и сестрам (¼)».
Отцовская тяга к многоженству пропадает, и он женит сыновей, если за один и тот же калым можно купить жену себе или три жены сыновьям, потому что 3 помноженное на ¼ дает ¾, а это больше, чем ½.
Генам, объясняет Бэрэш, которые сидят в этих необразованных африканских пастухах, не нужно быть доками в элементарной алгебре, потому что «эволюция уже произвела арифметические исчисления». Пилецкий делает эту глубокую мысль еще глубже, и произносит очередной великий афоризм про роль математики: «нам незачем постигать азы арифметики».
Конечно, каждый встанет на сторону Ломоносова, который считал, что «математику уже затем учить следует, что она ум в порядок приводит».
Все это напоминает пресловутого Дэникена, который в каждом стихе Иезекиля видит научно-сметную документацию летающей тарелки.
Один ген – один признак
ВВС
Гамильтона обычно критикуют за то, что он допускает мысль о существовании проекта сложного поведения в элементарной частице, которая кодирует синтез всего лишь одного белка. Докинз в этом случае справедливо говорит, что речь идет не о том, что «данный ген определяет некий признак», а о том, что «изменение этого гена вызывает изменение этого признака».
Есть, например, ген, который кодирует синтез белка р53. Этот белок, обнаружив потенциально опасную клетку, сдвигает равновесие в пользу апоптозных белков, и опасная клетка гибнет.
Если же в гене, кодирующем белок р53, происходит мутация, «система регуляции клеточного деления остается без должного присмотра» и злокачественные клетки начинают бесконтрольно расти. То есть, мутант р53 не борется с опасными для организма клетками, а наоборот своей халатностью способствует их росту.
Гипотетический ген альтруизма можно сравнить и с мутантным геном гемоглобина HbS. Нормальный ген гемоглобина HbA имеет в 6-й позиции бета-цепи глутаминовую кислоту. У мутанта в этой позиции стоит аминокислота валин, и это приводит к серповидноклеточной анемии.
У людей, страдающих этим заболеванием, гемоглобин имеет не овальную, а серповидную форму и практически не переносит кислород. Поэтому дети с генотипом гемоглобина HbSHbS не доживают до взрослого возраста.
Это значит, что за эгоистический можно принимать любой ген, участвующий в формировании сложного поведения, если только у него есть мутантный аллель, который меняет направление поведения на противоположное.*
Правда, эта модель возникновения альтруистического поведения, серьезно уступает дарвиновской в психологической достоверности.
По представлениям Дарвина все без исключения существа получают от природы качество, которое принято называть эгоизмом. Но есть избранные индивиды, которые помимо этого наследуют от своих родителей благородный характер. Если языком генетики, их генотип получает дополнительный ген, которого нет у эгоистов.
Почему бы Дарвину не предположить, что благородные наследуют благородный характер, а неблагородные – подлый?
Он не сделал этого предположения, потому как хорошо знал, что этот подлый элемент есть и в благородных натурах, что благородство и эгоизм имеют в индивиде одинаковые права на господство, и именно поэтому благородный индивид борется «между противоположными побуждениями». Благородство не всегда берет верх над эгоизмом, и тогда благородный человек испытывает «угрызение совести, раскаяние, сожаление и стыд».
Правда, сегодня нам известно, что организм не может наследовать дополнительный ген, и генотип одного человека не может быть меньше или больше, чем генотип другого.
Гипотеза Гамильтона предполагает, что человек есть что-то одно: или эгоист, или альтруист. В аппарате управления индивида есть только одна командная позиция, и она может быть занята только одной поведенческой программой, которую кодирует только один аллель: аллель эгоизма или аллель альтруизма.
Эта модель не может объяснить, как раз то главное, ради чего невинный Дарвин вводил в генотип человека дополнительный ген: почему человек борется «между противоположными побуждениями», что является самой характерной чертой нравственных переживаний.
С другой стороны, к принудительной силе альтруистического аллеля Гамильтон относится без особого энтузиазма и, по-моему, как-то не уверенно.
Можно подумать, что человек с альтруистическим генотипом по первому требованию бросится на помощь каждому, кто попал в беду, но сам Гамильтон, судя по всему, так не думает.
«Никто, - пишет он, - не готов пожертвовать своей жизнью ради одного человека, но он пожертвует, когда может спасти двух братьев, или четверых племянников, или восьмерых двоюродных братьев».
Получается, что к альтруизму организм принуждает не альтруистический генотип, а математический результат совокупной приспособленности.
h3> Совокупная приспособленность и популяционная ге
Совокупная приспособленность и популяционная генетика
Предположим, однажды у какого-то гена, который можно назвать геном эгоизма, произошла мутация, и в мир явился летальный альтруистический аллель (а). Потом этот аллель передался другому организму, и однажды они встретились и образовали альтруистический генотип (аа).
Гамильтон утверждает, что спасая индивида с аллелем альтруизма, альтруист спасает этот аллель от вымирания или от эллиминации.
Докинз поддерживает эту идею самым решительным образом.
«Если, - говорит он, - индивидуум умирает, чтобы спасти десять близких родственников, то одна копия гена, определяющего альтруизм может погибнуть, однако гораздо большее число копий того же гена будет спасено».
Но Гамильтон и Докинз глубоко ошибаются, когда думают, что аллель, спасенный, например, от хищника, спасен и от естественного отбора.
О чем на самом деле идет речь, когда альтруист спасает своего родственника с аллелем альтруизма?
Если проблему, поставленную Дарвином, изложить в генетических представлениях, можно сказать, что отбор будет уничтожать фенотипы с гомозиготами аа, что должно привести к вытеснению аллеля а аллелем А из популяции. Чтобы этого не происходило, альтруист должен обеспечивать численное превосходство аллелей альтруизма над аллелями эгоизма:
q ≥ p,
где q – частота аллеля альтруизма, p – частота аллеля эгоизма.
Может ли альтруист обеспечить такие условия для q?
Предположим, есть популяция, которая состоит из четырех братьев, рожденных родителями с генотипами Аа:
АА + 2Аа + аа.
Представим, что один, два или три брата альтруиста (АА, Аа, Аа) подвергаются смертельной опасности, и альтруист (аа) ценой собственной жизни спасает их от смерти.
АА + 2Аа – аа (1)
Рассчитаем частоты аллелей, которые останутся после альтруистического события по формуле
p = D + H или q = R +
H,
где p – частота аллеля А, q – частота аллеля а, D – частота генотипа АА, H – частота генотипа Аа, R – частота генотипа аа.
Подставив в эти формулы показатели выражения (1), получим, что частота аллеля А (p) равна – 0,75, а частота аллеля а (q) равна – 0,25.
Таким образом, вопреки предсказаниям Гамильтона, мы получаем выражение
q q1= q2(1–Sq)/(1–Sq2),
Это очень просто
объяснить.
Альтруист спасает от вымирания не аллель альтруизма (а), на что
надеется Гамильтон, а генотипы Аа и аа, которые содержат этот
аллель, и вместе с ним спасается от вымирания и аллель эгоизма
(А), который содержится в генотипах Аа и АА. При этом частота
аллеля а сокращается за счет затрат альтруиста, а частота аллеля
А остается постоянной, и эгоистический аллель, таким образом,
будет вытеснять из популяции аллель альтруизма (рис.1)
p1= p/(1–Sq2) > q1= q2(1–Sq)/(1–Sq2),
где S – коэффициент отбора равный единице (S = 1), если приспособленность аллеля а (W) равна нулю.
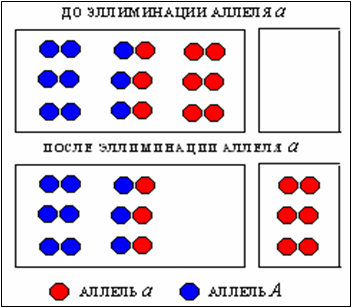
Здесь все происходит точно так же, как в случае с патологическим аллелем гемоглобина HbS , который вытесняется из популяции нормальным аллелем HbА.
С формальной точки зрения между вытеснением альтруистического аллеля и дефектного аллеля гемоглобина нет никакой разницы: в обоих случаях aa и HSHSпогибают, а Aa, AA и HAHS, HAHA не важно по какой причине, остаются в популяции.
По некоторым данным частота генотипа HbSHbS приблизительно 0,0000428 на одного человека, и это говорит о том, что аллель HbS элиминирует.
Правда, HbS имеет все еще высокую частоту, потому что в районах, где свирепствует малярия, отбор уничтожает генотипы HbSHbS и HbА HbА, а люди с генотипом HbАHbS имеют против малярии иммунитет.
Но это не меняет сути дела.
Так вот, частота аллеля а будет снижаться из поколения в поколение:
qt = , (2)
где qt – частота аллеля q через t поколений; q0 – начальная частота аллеля q; t – число поколений.
Рассчитывая время элиминации рецессивного аллеля при коэффициенте отбора равном единице (S = 1), по формуле (2) Айала и Кайгер получили следующие показатели (табл1):

Снижение частоты q при S = 1 Число поколений
От 0,99 до 0,50 1
От 0,50 до 0,10 8
От 0,10 до 0,01 90
От 0,01 до 0,001 900
От 0,001 до 0,0001 9000
Табл.1 Число поколений, необходимое для определенного снижения частоты аллеля (q) при коэффициенте отбора S = 1 против рецессивных гомозигот.
Если альтруизм, как считают, например, ученые из Киотского университета, известен шимпанзе, можно предположить, что человек и эти обезьяны должны были получить альтруистический аллель от своего общего предка. И поскольку линии шимпанзе и человека разделились 4-4,6 миллиона лет назад, получается, аллель альтруизма появился до этого события.
Из расчета, что у человека сменяются три поколения каждые сто лет, хоть так было и не всегда, можно посчитать, что со времени происхождения этого мифического аллеля до наших дней сменилось 120 000 поколений людей.
Аллель альтруизма, конечно, не смог бы исчезнуть за это время полностью. Сегодня его частота равнялась бы 0,000008 аллеля на человека. Но при такой частоте аллеля вероятность образования альтруистического генотипа равна нулю.
Эусоциальность и альтруизм
Всех особенно вдохновляют преимущества, которые открывает для совокупной приспособленности гаплодиплоидное определение пола.
Отец у гаплодиплоидных имеет в генотипе всего один аллель, и рабочие самки генетически отличаются друг от друга только аллелями, которые получили от матери. Поэтому среднее генетическое сходство (r) между сестрами равно ¾, а не ½, как было бы с собственными детьми. И рабочим выгоднее помогать матери, чем рожать собственных детей.
В самом деле, если не у всех гаплодиплоидных, то у эусоциальных матка, благодаря своей чудовищной плодовитости, вполне могла бы окупить любые жертвы своих добровольных помощников.
За 5-6 лет жизни царица производит около 3 миллионов стерильных самок, и 10-18 фертильных принцесс. Эти принцессы и есть тот выигрыш, который могла бы получить стерильная самка, воспитывая не детей, а сестер.
На первый взгляд, 18 принцесс на 3 миллиона стерильных особей, это бесконечно малая величина – 0,000006 на рабочую. Но каждая принцесса умножит эти 0,000006 на три миллиона новых рабочих, и получается, что неравенство Гамильтона R >c/b будет безукоризненно выполняться.
Среднестатистическая одиночная пчела может вырастить собственными силами 15,37 детей, если считать что Рыжая Осьмия (Osmia rufa) за сезон выхаживает по 24-40 детей, смолевщица (Anthidium septemdentatum) – 2, пчелы-каменщицы (Chalicodoma) – 6-15, листорезы (Megachile) – 17.
Как относятся затраты альтруиста к выигрышу родственников определим по формуле Крейга (Creig) и Вест-Эберхарда (West-Eberhhard)
(K =) > ,
где К – отношение выигрыша особи, пользующейся услугами альтруиста, к проигрышу альтруиста; ni – число потомков матки в расчете на одну рабочую; no – число репродуктивных потомков одиночной самки; r0 – генетическое сходство самки с собственным потомством; ri – генетическое сходство рабочей с репродуктивным потомством матки.
= 1,
17 ( ) =0,67
Этот результат будет еще лучше, если сделать расчеты для медоносной пчелы Скутеллата (Apis mellifera scutellata), которая обитает в Африке, роится 6-12 раз в год и за пять-шесть лет жизни производит на свет 270-324 принцессы.
Если к этому прибавить, что гаминтольянцы почему-то думают, что матка рожает исключительно особей с генотипом аа, то поневоле придет в голову, что рабочей выгоднее воспитывать чужих, чем рожать собственных детей.
Докинз даже говорит, что самка ухаживает «за своей матерью как за эффективной машиной, производящей сестер». «Рабочие, - пишет он, - используют свою мать как более эффективного производителя копий своих собственных генов», и «относятся к репродуктивным особям как к домашним животным».
Но фабрики массового производства альтруистических аллелей, это всего лишь очередная «Утопия» от Ричарда Докинза.
Во-первых, гаплодиплоидным сложно учредить такие фабрики.
Родители рабочих не могут иметь альтруистические генотипы, потому что по условиям Гамильтона они должны быть стерильны. Поэтому нужно сделать исключение, по крайней мере, для трутня с генотипом а. Вступая в брак с самкой, у которой генотип Аа он даст потомство с альтруистическим (аа)генотипом (табл.2).


А а
а Аа аа
Табл.2
Во-вторых, популяция эусоциальных насекомых с аллелем альтруизма все равно не смогла бы выжить. В такой популяции отбор стал бы тотально уничтожать не только альтруистический, но и эгоистический аллели.
Альтруистические аллели были бы уничтожены по понятной причине: согласно правилу Гамильтона они не оставляют потомков.
Эгоистические генотипы, произведенные генотипами АА, Аа и А обречены из-за массовой гибели колоний, поскольку все рабочие в таких колониях будут эгоистами, и царица будет испытывать недостаток рабочих рук и нести потери из-за ожесточенной борьбы между самками.
Слабыми и, скорее всего, нежизнеспособными были бы и колонии, образованные маткой с генотипом Аа и трутнем с генотипом а. Дело в том, что только половина их дочерей (табл.2) будет иметь альтруистический генотип (аа), и такая колония должна испытывать те же трудности, в которой все рабочие имеют генотипы Аа или АА.
Одним словом, альтруистический аллель мог бы привести эусоциальных насекомых к катастрофе.
Если предположить, что аллели а и А сохраняются в гетерозиготах и генотипы Аа и АА восстанавливаются, когда колония выходит из так называемого бутылочного горлышка (bottleneck), то после роения должна наблюдаться массовая гибель колоний, где среди рабочих преобладают особи с эгоистическими генотипами.
И как раз то, что таких событий в природе не происходит, говорит о том, что не существует и аллеля, который кодирует альтруизм у эусоциальных насекомых.
Нужна ли гипотеза Гамильтона?
Но прав ли был Дарвин и есть ли вообще необходимость в теории Гамильтона?
Дарвин рассуждал об этике не как натуралист, а как хорошо начитанный человек. О том, что альтруизм, это разновидность самоубийства, ему было известно от «многих очень талантливых писателей», которыми в то время «была обсуждаема эта проблема».
На самом деле достоверно известно только то, что альтруист рискует своей жизнью, но из этого вовсе не следует, что он ищет и находит смерть.
Если говорить только о человеке, то миллионы людей во всем мире систематически подвергают смертельной опасности собственную жизнь.
Американские пожарные, например, по статистике погибают на службе в три раза чаще, чем среднестатистический американский рабочий. C 1992 до 1997 на каждые 100 000 пожарных погибало 16,5 человек. Среди остальных профессий ежегодные потери на те же 100000 составляли 4,7 человека.
С 1994 по 204 год в США при исполнении служебных обязанностей от сердечных приступов, от травм и от удушья погибло 368 профессиональных пожарных и 610 добровольцев.
Традиционно-опасной считается профессия шахтера.
Первое упоминание о подземной катастрофе относится к 1376 году. Тогда на германском руднике «Rammelsberg» рядом с городом Гослар в результате обрушения кровли погибло больше ста человек. В 1448 году в Австрии затопило рудник «Heilig-Kreuz-Stollen» и погибло 260 человек. Самая масштабная промышленная авария XIX-го века произошла 12 декабря 1866 в английской угольной копи Окс в Барнсли. Тогда в результате взрыва погиб 361 человек (Гражданкин А.И., http).
Самая крупная катастрофа произошла в 1942 году на шахте «Honkeiko» в Манчжурии, где погибло сразу 1549 человек. В 1906 году во Франции на шахте «Courrières» во время аварии погибло 1090 шахтеров. Крупнейшая в России шахтная авария произошла в 1908 году на шахте № 4-4 бис Макарьевского рудника в Юзовке. Тогда погиб 271 горняк.
В Великобритании с 1708 по 1966 год в 66 угольных катастрофах погибло 8462 человека. Число погибших в авариях на угольных шахтах Великобритании в XVIII - ХХ вв. с 1700 по 2000 гг. 15 068 человек. В США с1839 г по 1972 года в 37 крупных авариях погибли 5269 человек. В Германии с 1867 по 1962 год погибли 3144 человека.
В границах бывшего СССР за последние 120 лет (1890-2010 гг.) только в особо крупных, крупных и больших шахтных авариях смертельно травмированы 1983 человека.
Вообще говоря, риск, это не что-то исключительное, крайне редкое, несовместимое с жизнью, это повседневное и повсеместное явление.
Найт показывает, «что именно риск является основой предпринимательской прибыли» или предпринимательского поведения.
Самым выразительным примером предпринимательского риска может служить венчурный бизнес. Термин venture обозначает предприятие, связанное со значительным риском. Венчурный бизнес типичен для коммерциализации результатов научных исследований в наукоемких областях, где нет никаких гарантий получить прибыль.
Что такое риск?
Однако важно здесь то, что в условиях смертельной опасности работают миллионы людей, а погибают единицы.
Если говорить только о шахтерах, то, например, в 80-е годы XIX-го века в горнозаводской промышленности Российской империи среднегодовой уровень смертельного травматизма составлял 0,53±0,05 погибших на 1 тысяч горнорабочих.
По статистике, которую приводит Теодор Фридгут, ежегодно в угольных шахтах в конце девятнадцатого начале двадцатого веков в разных странах мира гибло от 0,98 до 3,60 человека на каждую тысячу забойщиков.
Этимологически, рисковать, значит лавировать. Не идти на верную смерть, а избегать смерти, уворачиваться от опасности. И было бы правильнее сказать, что риск, это какое-то искусство, какая-то ловкость, какая-то расчетливость, которая делает животное неуязвимым перед смертью.
И хотя поражение в предпринимательской деятельности в большинстве случаев не заканчивается трагически, законы риска, которые действуют в экономике, могут объяснить природу риска и в других отраслях социального поведения.
Найт проводит различие между риском и неопределенностью, и говорит, что риск, это некое количество, доступное измерению. По его словам, это «измеримая неопределенность».
Попросту говоря, риск опирается на предвидение благополучного исхода событий. Это предвидение возможно «либо благодаря априорным расчетам, либо из статистических данных прошлого опыта».
Идеальным воплощением расчета в рискованном поведении может служить технология добычи каменного угля и правила техники безопасности при работе в забое.
По статистике, которую приводит Либерман, шансы шахтера выжить в забое увеличиваются в зависимости от стажа и продолжительности работы. 70,5 процентов жертв в угольных забоях работали на шахте шесть месяцев или менее, еще 15,9 процентов имели год опыта или менее, и только 13,6 процентов работали более одного года.
Из произвольно взятых Либерманом 63 случаев гибели в шахтах, 11 вызвано нарушениями правил добычи, 34 произошло из-за недостатка навыка или внимания. Остальные случились по вине работодателей.
Для чего нужен риск?
Но что заставляет человека рисковать, если всегда есть альтернатива поберечься от опасного предприятия?
Традиционно риск связывают с возможностью ущерба, и, во всяком случае, в этике именно этот ущерб рассматривают, как добродетель. Так же и предпринимательский риск в классической теории отождествляют с математическим ожиданием потерь.
Это представление не дает риску никакого разумного объяснения.
И только в неоклассической теории у А. Маршалла и А. Пигу, риск предполагает «не только опасность не достичь намеченного результата, но и вероятность превысить ожидаемую прибыль».
Собственно говоря, эта-то вероятность получить сверхприбыль и заставляет человека рисковать, не считаясь с опасностью потерять стабильный и безопасный доход. Человек всегда рискует там, где есть приз, где есть какая-то бо′льшая выгода, какое-то бо′льшее преимущество.
Моббинг
Примером, когда животное получает эволюционное преимущество, подвергая смертельной опасности собственную жизнь, может служить моббинг.
Спангенберг в своих замечательных записках рассказывает о воробышке, который жил у них дома. Этот воробей обыкновенно сидел на углу стола, заглядывал в глаза матери натуралиста и чирикал, выпрашивая поесть. Мать его кормила, и птенец испытывал к ней сильную привязанность.
Еще в доме у Спангенбергов жил старый охотничий пес по кличке Маркиз. Он тоже был привязан к хозяйке дома, и из-за этого попал в немилость воробью. Однажды пес, как обычно, подошел и положил морду на колени хозяйке, и тут разгневанный воробьишко воинственно бросился ему на голову.
Пес стряхнул воробья на пол, издали и с опаской его обнюхал, и не решился наказать наглеца. Этот случай, пишет Спангенберг, не прошел для Маркиза бесследно. После этого происшествия «он не решался подходить к хозяйке, когда около нее вертелся воробей-забияка».
Это классический образец моббинга, поведения, когда не хищник нападает на жертву, а жертва на хищника, и когда, художественно говоря, смерть вынуждена отступить там, где она просто неизбежна.
Другой хрестоматийный пример моббинга показывает пеганка (Tadorna tadorna). О пеганках, пишет Лоренц, «давно известно, что они люто травят хищников; их яркое оперение настолько угнетает лис, что они могут безнаказанно высиживать утят в лисьих норах, в присутствии хозяев».
Болгары поэтому называют пеганку «лисьим пещерным гусем»
«Лисицы и пеганки, - пишет Боинус, - живут в одной и той же норе; первая, вообще не щадящая никакой другой птицы, не обижает пеганки. Причина такого странного отношения разбойницы-лисицы к пеганке заключается, вероятно, в том, что лисе внушает почтение необыкновенная храбрость этого гуся».
Боинус наблюдал, например, как совсем маленькие птенцы пеганки «оборонялись от нападения крупных домашних птиц и других животных».
Однако он же говорит, что это сожительство пеганки с лисой «оказывается не вполне безопасным». Ему пришлось найти около одной лисьей норы крылья и перья пеганки, хотя он и не уверен, что утку съела именно квартирная хозяйка.
И пеганки – не единственная птица, которая может жить в одной норе с хищником. Такое соседство встречается иногда и у огарей.
Судя по всему, моббинг выполняет ту же функцию, что и охрана территории. Исследовательская группа экосистем из университета Нового Брунсвика проводила свои исследования на синицах-гаичках, которые только в 3 случаях из 37 участвовали в моббинге вне границ своих территорий.
Моббинг считают групповым поведением, и можно подумать, что в одиночку животное никогда не решится нападать на хищника. На самом деле между животными, которые травят хищника, не может быть предварительной договоренности, и они нападают на хищника именно в одиночку.
Во всяком случае, в экспериментах Roni Ostreiher, арабские говорушки нападают на рогатую гадюку и группой и поодиночке, и эти поединки, описанные по нескольким параметрам, мало чем друг от друга отличаются.
В одиночку говорушки продолжали поединок 6,45 ± 1,7 минуты, группой – 8,2 ± 1,5. Вместе говорушки были несколько смелей, чем в одиночку, и приближались к змее на 2,5 сантиметра ближе, чем порознь.
Можно предположить, что говорушки, вступая в поединок с гадюкой, рассчитывают, что на их крик слетятся сородичи. Но из экспериментов Roni Ostreiher напрашивается вывод, что сами драчуны не рассчитывают на постороннюю помощь. Количество сигналов, которые подают говорушки, нападая на змею в одиночку, на 3,2 меньше, чем та же говорушка подает, нападая на хищника в компании.
Леандро де Кастро-Сикира рассказывает о случае, когда в жилом районе Бразилии на лесного козодоя напали две мухоловки, три рыжих Fernerius и семь белобровых пересмешников, которые, конечно, не могли договориться между собой.
Просто сородичи и птицы другого вида всегда оказываются рядом и слетаются на шум, как это вообще принято у многих птиц, если кто-нибудь затевает драку с хищником или между собой.
Мобинг у животных, как и любое другое рискованное поведение, опирается на расчет. Животные, нападая на хищников, соизмеряют свои силы и преимущества.
На сурикатов, например, охотится много самых разных хищников. Это хищные птицы, млекопитающие, такие, как шакалы и дикие африканские кошки, и змеи. Завидев хищную птицу, сурикаты прячутся в норы. От наземных хищников они или спасаются бегством, или травят их.
Встречи со змеями, по словам Beke Graw и Marta B. Manser, почти всегда приводят к моббингу, хотя «мобинг змей чаще всего заканчивается бегством сурикатов». В 88 случаях из 100 сурикаты травят полосатых хорьков, в 67 случаях – черепах и в 61 случае желтых мангустов.
Эта разница в вероятности моббинга разных животных как раз и указывает на то, что, нападая на хищников, сурикаты проявляют не безрассудную храбрость, а опираются на какой-то эмпирические соображения.
Альтруисты Дарвина
Я думаю, если бы Дарвин подошел к проблеме альтруизма не как философ-идеалист, а как натуралист, он бы сделал вывод, что альтруизм не препятствует, а наоборот способствует борьбе за существование.
Вот портрет альтруиста, описанный Дарвином со слов «заслуживающего доверия писателя», который рассказывал о бойцовых петухах (Gallns Stanleyi).
«Однажды, - рассказывает он, - ястреб бросился на бойцовую курицу, сопровождаемую цыплятами; вдруг петух прибежал на выручку и пронзил шпорой глаз и череп хищника».
Вообще, петухи рода Gallns, замечает великий натуралист, обладают «неукротимым мужеством», или благородным характером. И он, между прочим, пишет, что родственный Gallns Stanleyi дикий вид Gallns Stanleyi тоже отчаянно дерется, «защищая свой гарем».
Здесь Дарвин описывает существо, которое не отступает перед смертельной опасностью, защищая других существ, и имеет целый гарем, целую фабрику, которая производит наследников его благородного характера.
Само по себе владение гаремом указывает так же и на то, что альтруисты занимают высокое положение в иерархии, что для «наиболее мужественных» совсем не удивительно.
И игра со смертью не мешает этим деспотам и героям дожить до почтенной старости. Вот свидетельство самого Дарвина, сделанное со слов Брэма, наблюдавшего павианов в Абиссинии.
Большая толпа павианов, пишет он, переходила через долину. И когда одни уже взобрались на гору, а другие еще оставались внизу, на них напали собаки. Тогда «старые самцы тотчас бросились вниз со скалы, широко раскрыв пасти и с
таким страшным ревом, что собаки быстро отступили».
Брэм ясно указывает, а Дарвин за ним повторяет, что эти альтруисты были уже старыми.
В самом деле, это просто нелепо предполагать, что существо, способное успешно бороться за других, проигрывает борьбу за существование тем, кто в опасную минуту рассчитывает на его помощь.
Все, одним словом, говорит за то, что риск, не обезоруживает индивида перед естественным отбором, а, наоборот, способствует его выживанию. Способность рисковать, судя по всему, это показатель приспособленности, и он не имеет ничего общего с безрассудным самоубийством.
Есть ли ген альтруизма?
Гамильтон, излагая свои идеи, основанные на мифическом гене альтруизма, не знал ничего определенного о его существовании. И сегодня нет никаких убедительных доказательств, что такой ген действительно существует.
Карин Вервей (Karin J.H. Verweij) со своими коллегами сравнила личностные качества 5117 человек с 1 252 387 генетическими маркерами и не один маркер не указал на какую-нибудь связь с чертами личности.
Она, конечно, искала связь между генетическими маркерами и такими качествами, как осторожность, любопытство, честолюбие и настойчивость, и это только косвенно указывает на то, что и поиски гена альтруизма тоже не увенчаются успехом.
Правда, несколько лет назад начали появляться сообщения об открытии этого пресловутого гена. Первопроходцем стал профессор Иерусалимского университета Ричарда Эбштейн.
Эбштейн на роль гепотетического гена выдвигал снип гена окситоцина OXTR rs1042778 и в меньшей степени снип rs237887 и ген рецептора вазопрессина AVPR1a.
Однако шведский генетик Дэвид Кесарини (David Cesarini) в своих экспериментах не подтвердил результаты, полученные Эбштейном в экспериментах со снипами окситоцина. «Мы, - пишет он, - не нашли убедительных доказательств, что снипы
rs1042778 или rs237887 являются источниками индивидуальных различий игроков в игре Диктатор».
На работу с геном рецептора вазопрессина Эбштейна вдохновила «замечательная серия исследований» Янга.
Ларри Янг (Larry J. Young), профессор кафедры психиатрии и поведенческих наук университета Эмори в 1999 году в журнале Hormones and Behavior опубликовал статью, в которой утверждал, что рецептор AVP V1a программирует моногамное поведение у прерийной и сосновой полевок.
«Моногамные прерийные (Microtus ochrogaster) и сосновая (Microtus pinetorum) полевки, - пишет он, - имеют аналогичный OTR и рецептор V1a, которые не совпадают с соответствующими рецепторами немоногамных горных (Microtus montanus) и луговых (Microtus pennsylvanicus) полевок. Эти специфические различия в рецепторах содействуют различиям в поведении».
Правда, группа Джеральда Хекеля (Gerald Heckel) из лаборатории молекулярной генетики Бернского университета не подтвердила выводов Янга.
Группа Хекеля проверила предсказания Янга, обследовав на предмет наличия или отсутствия avpr1a STRs у полевок, собранных с трех континентов – из Европы, Северной Америки и Азии. Они выявили выявил наличие avpr1a STRs у 19 видов Microtus из 21, а также у трех других грызунов, родов и семейств (полевки, Clethrionomys glareolus, водяная полевка, Arvicola terrestris; и домашние мыши, Mus musculus).
«Наши результаты, - пишет Хекель, - по разделению avpr1a STRs в эволюции рода Microtus и других млекопитающих опровергают связь между наличием этих генетических элементов и моногамией у грызунов и других таксонов».
Тем не менее, Эбштейн с энтузиазмом взялся за дело.
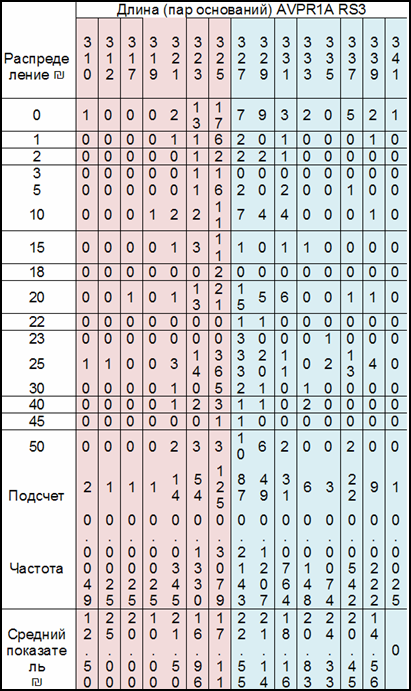
В его эксперисенте участвовало 203 человека (102 мужчины и 101 женщина). Они играли в игру «Диктатор». Игрок получал пятьдесят шекелей и на свое усмотрение мог поделиться деньгами с анонимным партнером или оставить всю сумму себе.
Каждый игрок был так же протестирован по гену AVPR1a. Этот ген имеет две формы. Одна форма отличается от другой длиной промоутера, и для удобства принято называть одну форму короткой (S = 308-325bp), другую – длинной (L = 327-343bp).
Все игроки распределись по генотипам следующим образом: SS – 24 процентов игроков; SL – 48 процентов; LL – 27 процентов.
Самыми щедрыми оказались персонажи с генотипом LL. (Табл.3).

Таблица 3. Среди персонажей с генотипом LL высокие отчисления партнерам сделали 61,8 процента игроков, низкие – 38,1 процент; у индивидов с генотипом SS высокие отчисления сделали 26 процентов игроков, низкие – 74; у игроков с генотипом SL эти показатели разделились примерно поровну – 51,0 и 48,9 процентов.
ААнализ результатов Эбштейна
Почему Эбштейн решил, что длинный аллель гена AVPR1A, это и есть ген альтруизма?
Выборка, которую он сделал, показывает, что среди персонажей с длинным аллелем в доминирующей позиции генотипа, преобладают те, кто пожертвовал анонимному получателю больше 25 шекелей, а тех, кто пожертвовал меньшую сумму составляют меньшинство. Среди игроков с коротким аллелем было больше тех, кто отдал партнеру меньше 25 шекелей, и меньше тех, кто отдал больше 25 шекелей (табл. 4).
Другими словами: с анонимными партнерами
делились все, но среди игроков с длинным аллелем было больше щедрых,
среди игроков с коротким аллелем было больше скупых (рис. 2)
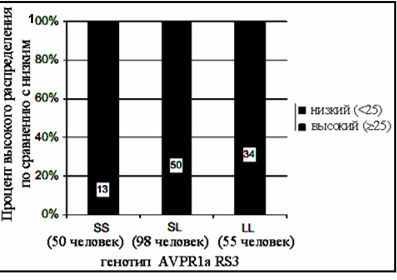
Из этого Эбштейн и делает вывод, что аллель AVPR1A с парами оснований от 327 до 343bp и есть гипотетический ген альтруизма. Его нисколько не смутило, что не все его альтруисты готовы делиться с партнерами, а эгоисты, наоборот, делают это иногда с большей охотой, чем альтруисты. Это очевидное недоразумение он объясняет влиянием среды.
Речь, одним словом, идет о норме реакции генотипа, о том, что генотип определяет не фенотип, а «скорее спектр возможных фенотипов».
Но когда, например, Айала и Кайгер говорят о норме реакции генотипа, они имеют в виду совсем не тот генотип, о котором думает Эбштейн. Речь идет не о комбинации аллелей одного гена, а о совокупности всех генов организма.
И, следовательно, объяснить, почему эгоистический генотип у Эбштейна дает альтруистический результат, и, наоборот, альтруистический генотип – эгоистический можно только тем, что ни тот ни другой не связаны с щедростью или скупостью игроков.
Ведь и у Эбштейна средний показатель щедрости игроков показывает, что все они одинаково щедры и одинаково скупы. Индивиды, у которых в доминантной позиции генотипа стоит длинный аллель, в среднем отдавали по 17,75 шекелей против 17,58, которые отдавали индивиды с коротким аллелем и были щедрее только на 0,17 шекелей. Это всего 0,34 процента (табл. 5).
12.5
310 312 317 319 321 323 325 327 329 331 333 335 337 339 341
0 25.00 20.00 10.00 21.50 16.96 17.11 22.55 21.14 18.16 20.83 24.33 20.45 14.56 0
Табл. 5
Эта разница просто вопиюще мала, чтобы быть причиной той пропасти, которая, как многие думают, существует между альтруизмом и эгоизмом.
Что такое альтруизм?
Мне кажется лучше всех наше представление об альтруизме выразил биолог и апологет Гамильтона в России доктор Марков. Он говорит: всем ясно, что такое альтруизм.
На самом деле, это-то как раз и есть самое неясное место во всех рассуждениях об альтруизме.
Все говорят, что быть альтруистом, значит жить для других, но всем так же хорошо известно, что никто не живет для других и за фасадом бескорыстия каждый раз обнаруживается эгоистический мотив.
Очень интересная подробность: Гамильтон, по словам Шварца, не был убежденным сторонником идеи альтруизма.
Он считал, что «истинное бескорыстие не встречается почти никогда», и мог назвать только двух людей, которые могли претендовать на звание альтруиста. Это была мать Тереза и друг Гамильтона Джорж Прайс, который подвел под теорию группового отбора свою математическую основу, посчитав выкладки Гамильтона слишком громоздкими.
Если считать, что заслуга Гамильтона в науке состоит в том, что он подвел математическую базу под идею, высказанную впервые Ходлейном, то станет понятным почему в этот короткий список попал Прайс.
Прайс подвел под теорию семейного альтруизма свою математическую основу, посчитав выкладки Гамильтона слишком громоздкими, и вместо того, чтобы оспаривать свое первенство, занялся толкованием Библии и помогал материально лондонским алкоголикам.
Другим человеком, который мог бы подтвердить альтруистический характер Прайса был Джон Мэйнард Смит. В 1968 году Прайс впервые предложил рассматривать конфликты между животными с позиции теории игр. Он написал об этом статью, но не стал ее публиковать.
Рецензировал эту статью Джон Мэйнард Смит. Он сумел оценить идею Прайса и в 1972 года опубликовал в Nature статью «Логика конфликтов в животном мире», положив, таким образом, начало применению теории игр в биологии.
И Гамильтон, и Смит оказались порядочными людьми. Гамильтон никогда не скрывал той роли, которую в его научной карьере сыграл Джорж Прайс. А Смит свою первую статью о теории игр подписал своим именем и именем Прайса.
Одним словом, у Гамильтона были все основания считать Прайса чуть ли не единственным в мире альтруистом. Но есть что-то очень важное в том, что в 1975 году Прайс перерезал себе горло.
Это случилось после того, как он вернулся из загородного дома Гамильтона, где он гостил больше недели, и где, по словам Шварца к нему вернулся интерес к науке. И, тогда, когда о Гамильтоне заговорил весь научный мир.
Что касается Матери Терезы, то она была образцом христианского милосердия, которое, как и любое другое милосердие, опирается на эгоистическую природу.
Мир по христианской версии, это мир товарных отношений. Христиане знают преступление, за которым приходит возмездие. Они знают даяние, за которым должно последовать воздаяние.
«Давайте, и дастся вам, - говорит Иисус. - какою мерою мерите, такою же отмерится и вам» Павел пишет, «кто сеет скупо, тот скупо и пожнет». И Мать Тереза говорит: «чем больше ты разделишь с другими, тем большим будешь обладать».
Иисус, вот адресат, который получает благодеяния этой великой бессребреницы. И всё, и все были для нее олицетворением Иисуса, которому она хотела помочь: голодный, жаждущий, бездомный, больной, одинокий, нежеланный, прокаженный, нищий, слепой, калека, заключенный. Поэтому Иисус у Терезы, в том числе «человек с зачерствевшим сердцем, которому надо помочь его смягчить».
Правда, и среди христиан Мать Тереза была скорее исключением чем правилом. Когда она умерла, в ее ордене работало 4000 миссионеров. Когда она начинала свое предприятие, это был «дерзкий, не обеспеченные материально и предпринятый в одиночку крестовый поход против бедности городских кварталов».
Ясно, что принцип жить для других не определяет альтруизм как самостоятельную сущность. Это бросается в глаза, когда альтруизм путают с благотворительностью, как, например, делают Эбштейн и Христиан Монтаг (Christian Montag), пытаясь отыскать ген альтруизма.
Щедрость, которую Эбштейн и Христиан Монтаг, рассматривают как меру альтруизма, это в действительности мера неравноценного обмена, который еще не имеет представления о стоимости труда, и труд или услуги вознаграждаются не по их рыночной стоимости, а по усмотрению работодатели или получателя услуг.
С античной эпохи и до наших дней благотворительность была способом превращения богатства в политическое влияние. Благодетели заявляют своими благодеяниями, что им «можно передать бразды правления».
В республиканском Риме эта особенность благотворительности превратилась в серьезную политическую проблему, с которой пытались бороться законодательно.
В 181 первом году до новой эры в Риме «по предложению Гая Орхия был принят закон об обедах, ограничивший число гостей, которых гражданин Рима мог собирать за своим столом.
Двадцать лет спустя «был принят более строгий Фанниев закон». Он ограничивал «расходы на пиры 100 ассами в праздники и 10 ассами в обычные дни, а число гостей тремя-пятью человеками».
Наконец, в 143 году до нашей эры был принят закон Тита Дидия. «Он распространял действие закона Фанния на территорию всей Италии и привлекал к ответственности не только устроителей пира, но и гостей».
«Римляне, - говорит по этому поводу Монтескье, - своими законами противодействовали некоторым слабостям самой прочной в мире власти – власти добродетели».
По словам Транквилла, Цезарь домогался сана великого понтифика «с помощью самой расточительной щедрости». Саллюстий говорит, что Цезарь «достиг славы, одаривая, помогая, прощая».
Во время четвертого триумфа Цезарь потратил 65 тысяч талантов. Только для угощения народа накрыли 22 тысячи столов.
Римляне, - пишет Утченко, были падки на подаяния во время триумфов полководцев и щедрот политиков, «завоевывавших себе таким путем и популярность и голоса».
Даяние не утрачивает свой эгоистический мотив даже тогда, когда благотворитель отдает все, что у него есть.
Примером может служить потлач – взаимный обмен подарками, который продолжается до тех пор, пока дарители не становятся нищими.
В потлаче, по словам Мосса, «прежде всего отразилась борьба знати между собой за место в иерархии». Утверждение власти, таким образом, это и есть причина, эгоистический мотив разорительного дарения в потлаче. Даритель получает власть над получателем подарка, и о вожде говорится, что он «проглотил племена», которым раздал свои богатства.
Разница между простой благотворительностью и потлачем состоит только в том, что благотворитель хочет купить своими благодеяниями то, что даритель на потлаче демонстрирует как свою собственность.
Наконец, и смертельная опасность, которой подвергает себя индивид, спасая другого индивида от смерти, не устраняет корыстный мотив поступка, потому что альтруист повышает свою приспособленность.
Везде, одним словом, на месте, которое в нашей культуре приготовлено для мифической бескорыстной деятельности, сидит самый заурядный эгоизм.
Избавиться от этого навязчивого персонажа не сумел даже Кант, единственный, по-моему, моралист, который специально занимался этой работой.
Он со своим немецким педантизмом самым тщательным образом удалил из поступков корыстный мотив и вывел свой категорический императив: «поступай лишь по правилу, относительно которого ты вместе с тем можешь желать, чтобы оно имело силу всеобщего закона для всех разумных существ».
Но Шопенгауэр, такой же педант немецкого образца, тут же нашел, что и «здесь, несмотря на возвышенные априорные прелюдии, на судейском кресле восседает и вершит дела собственно эгоизм».
«Содержащееся в верховном правиле Канта указание на способ отыскать подлинный моральный принцип, - пишет он, - основано именно на молчаливом предположении, что я могу желать лишь того, что для меня всего выгоднее».
Однако при том, что в любых поступках мы без труда находим эгоистический мотив, человек почему-то легко отличает поступки альтруистические от эгоистических.
Наше этическое чутье объясняется тем, что противоположность между эгоизмом и альтруизмом состоит не в цели, которую преследует индивид, а в средствах достижения этой цели. Не в том, к чему он стремится, а в том, как он это делает.
Разница состоит в том, что тот, кого называют альтруистом, рискует потерять все, чтобы приобрести, а тот, кого называют эгоистом, приобретает все, ничем не рискуя.
Двоякая природа альтруизма
Как человек может жить для других, живя для себя? Каким образом возникает альтруистический эффект, если альтруист заботится о своей выгоде?
Обычно альтруистическое поведение, которым движет эгоистический мотив, представляют как инвестирование в выгодный проект. По Гольбаху «чувствующее и разумное существо» нуждается в чужой помощи и может ее получить, если само будет помогать другим.
Триверс, между прочим, в этом очень сильно сомневается, и, по-моему, он прав. У эгоиста не может быть никакой уверенности, что банк, в который он вкладывает, никогда его не обманет.
На самом деле альтруистический эффект возникает не как результат воли или внутренней потребности альтруиста, а как характерная особенность общественного образа жизни.
Когда говорят, что альтруист живет для других, то подразумевают, что он живет интересами других людей. Это можно понять, как то, что интересы альтруиста не совпадают с их интересами, и, если он будет преследовать свои интересы, то не сможет ублажить чужие.
На самом деле чужие интересы, это общие для всех, типичные индивидуальные интересы членов одной социальной группы. Именно благодаря этим типичным индивидуальным интересам создается общественное благо.
Известно, например, что термитник для термитов, это их общее благо. Но когда Грассе попытался понять, «у кого находятся чертежи гнезда» африканских термитов из рода кубитермес (Cubitermes) и макротермес (Macrotermes), он пришел к выводу, что эти практически одинаковые чертежи находятся у каждого термита. И каждый термит в каком-то смысле даже не подозревает о существовании других термитов.
И можно сказать, что термит создает общественное благо не потому, что в этом состоит его общественная идея, а потому что он преследует свои индивидуальные интересы.
Для иллюстрации альтруизма у человека Дарвин выбрал войну между племенами. Война, каждый сказал бы: к сожалению, самая подходящая стихия для проявления альтруизма.
Так вот, на поле боя альтруист встречается со своим личным врагом.
Это, конечно, общий враг племени или народа, но этот враг потому и называется общим, что он личный враг для каждого члена племени. Этот враг пришел убить всех, и альтруиста в том числе, и поэтому альтруист преследует личный, эгоистический интерес: победить врага и выжить, а победив, он избавляет от опасности тех, кто мог бы стать жертвой их общего врага.
Таким образом, альтруистический эффект возникает тогда, когда индивид, стремясь к максимальной собственной выгоде, максимально увеличивает общественное благо.
Юнг-Кью Чой и Сэмюэль Боулз (Jung-Kyoo Choi и Samuel Bowles) называют альтруизм на войне общинным (parochial altruists), и считают, что человеческий альтруизм развился из parochial altruists.
В этом прилагательном parochial у них содержится указание, если не на порочную, то на испорченную, злобную природу военного альтруизма.
Смущение, и не только у них, вызывает враждебность альтруиста к чужакам, потому что она не вяжется с любовью к человеку, которую традиционно приписывают альтруисту. Поэтому Чой и Боулз рассматривают parochial altruists, как частный случай альтруизма, как еще недоразвитый альтруизм.
На самом деле, чужаки выступают здесь не как олицетворение человека вообще, которого нужно любить, а как фактор опасности, и враждебность к ним альтруиста, это частный случай его естественной враждебности к любой опасности, от которой он защищает других индивидов.
И то, что альтруист не может оказать альтруистическую помощь чужим, говорит о том, что он не может этого сделать, когда у него нет личной заинтересованности.
Выводы
Таким образом я считаю, что теория альтруизма Уильяма Гамильтона построена на неверном представлении о генетической реальности и неверном представлении об альтруизме.

Сайт создан в системе uCoz |
Сайт создан в системе uCoz
|
|
