|
Максимальная медопродуктивность пчелиной семьи зависит от количества выращенного рабочими особями жизнеспособного расплода к периоду главного медосбора.
Родоначальницей в каждой семье пчёл является плодная матка, которая живет несколько лет только в составе семьи и ежегодно в период цветения медоносов откладывает в ячейки сот своего гнезда множество яиц оплодотворенных сперматозоидами трутней, из которых рабочие пчёлы воспитывают многотысячное потомство женских рабочих особей-сборщиц кормовых запасов из нектара и пыльцы.
Сбор кормовых запасов в каждой пчелиной семье зависит в первую очередь от наличия: наибольшего числа сборщиц; достаточной местной кормовой базы медоносов; условий внешней среды; наследственных качеств потомства полученного от родителей в естественном отборе. В отборе, как правило, выживали только те семьи, которые приспособились к окружающей их среде обитания.
Связь между родителями и потомством проявляется в информационной наследственности, которая передается от одного поколения другому через гены расположенные в определенной последовательности в хромосомах каждой особи. В одной хромосоме каждый ген занимает определенное место и контролирует один признак или несколько признаков наследственности. Известно, что все живые существа состоят из клеток, а в ядре клетки находятся хромотиновые зёрна, из которых путем деления образуются хромосомы. В каждой клетке любого организма у различных видов животных содержится, как правило, определенное число хромосом. По числу хромосом определяется пол животного или насекомого. У медоносной пчелы пол определяется с развитием рабочих пчёл и маток из оплодотворенных яиц, а трутней из неоплодотворенных яиц. Эту закономерность впервые открыл великий силезский пчеловод Ян Дзержон в 1845 году (Шаскольский, 1968).
Клетки организма (соматические) женских особей пчёл имеют диплоидный набор хромосом (2x16=32), то есть двойной. Соматические клетки мужских особей (трутней) содержат гаплоидный набор, т.е. они происходят из гамет матки и в своих клетках тела имеют по 16 хромосом (n - один геном). В половых же органах матки и пчелы-трутовки развитые яйца в яичниках содержат по 16 хромосом. В семяприемнике плодной матки и в половых органах половозрелого трутня, сперматозоиды содержат по 16 хромосом (рис. 6).
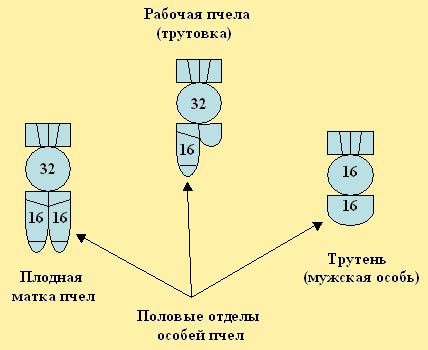
Рис. 6. Число хромосом в клетках пчёл
В результате из оплодотворенных яиц развиваются диплоидные женские особи-матки и рабочие пчёлы, а из неоплодотворенных яйцеклеток (партеногенез) развиваются мужские особи трутни (рис. 7), имеющие гаплоидный (одинарный) набор хромосом.
Иногда, из неоплодотворенного яйца может развиться диплоидная особь с 32 хромосомами. Такие случаи можно наблюдать у осиротевшей длительное время семьи пчел. Из яиц пчёл трутовок, пчёлы выводят матку неспособную к оплодотворению трутнями.
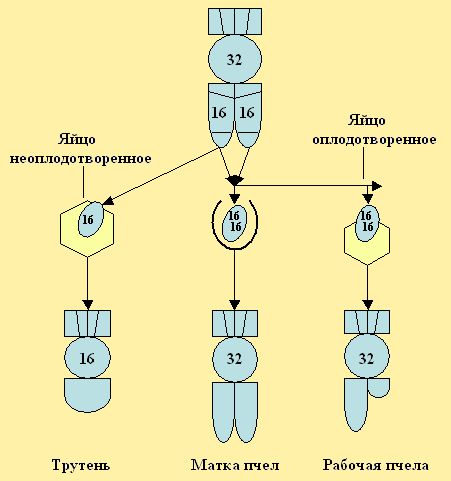
Рис. 7. При откладке, плодной маткой , в ячейки сот, оплодотворенных яиц, развиваются рабочие пчёлы и матки, их тело имеет 32 хромосомы, трутень - из неоплодотворенного яйца - 16.
Гены, хромосомы, аллели и мутации
В хромосомах распложены гены, которые содержат наследственную информацию. Следовательно, ген является единицей наследственной информации. Ген определяет развитие у пчёл одного или нескольких признаков. Число генов во много раз больше, чем хромосом. Хромосома передается по наследству как единое целое при коньюгации (процесс - сближения) и расхождение в мейозе, сохраняя свою целостность, а потому все гены, локализованные в одной хромосоме, обычно наследуются совместно. У пчел, каждая хромосома содержит один из аллелей гена.
Гены взаимоисключающих признаков составляют пару аллелей. Например, признак желтой окраски и темной (серой) окраски или признаки злобности и миролюбия у рабочих пчел, такие признаки в мейозе оказываются в разных гаметах.
Разные аллельные формы возникают при мутации (изменение наследственного материала) гена, свойственного дикому (нормальному) типу, где в паре аллелей один ген нормальный, т. е. неизменный, а другой измененный, то есть мутантный. Мутации, одного аллеля, нормального гена могут возникать несколько раз, в результате образуется серия аллелей. Это явление называют множественным аллелизмом - т. е. более двух аллельных форм гена. Ряд мутаций могут находиться в разных доминантно-рецессивных отношениях друг к другу и могут вызывать появления серии аллелей (C1, C2, C3,...Cn, которые составляют ряд. Мутации по значению для организма пчёл могут быть полезными, вредными, нетральными, а по характеру возникновения - естественные или при искусственном оплодотворении.
Для обозначения аллелей в генетике принята буквенная символика, в которой доминантный признак (признак проявляется у гибридных особей подавляя внешней рецессивный) обозначают заглавной буквой А, В или С и т.д. Рецессивные признаки (внешне скрытые) обозначаются строчными буквами - а, в, с и т. д. Если, доминантный признак не полностью подавляет рецессивный, то такое явление называют неполным доминированием.
У медоносных пчёл пол определяет не вся хромосома, а только её участок
- ген, он является элементарной единицей наследственной информации.
О. Маккензен и Робертс с 1951 года (Шаскольский, 1966) обозначили ген пола у пчелы, как и у наездника хабробракона (из семейства наездников браконид), через Х, а его аллели через Ха, Хb, Xc...... Для простоты аллели принято обозначать, как а, в, с, и т.д., помня, что это аллели гена Х (Шаскольский, 1966).
Известно, что каждый ген имеет множество различных типов: а, а2, а3,...а6......а12, которые называют половыми аллелями. Если в оплодотворенном яйце встречаются 2 аллеля, например, как а2а8 или а5а11 всегда разовьется самка, а если - а2а2 или а8а8 - разовьется самец, но он будет для пчёл нежизнеспособным, как правило пчёлы такие яйца поедают. Если рабочая пчела или матка имеют два одинаковых аллеля, например, АА или аа т.е. гомологичную пару, то они будут называться гомозиготными по паре одинаковых аллелей, а если аллели различны, как Аа, Вв, Сс и т.д. то такие особи называют гетерозиготными.
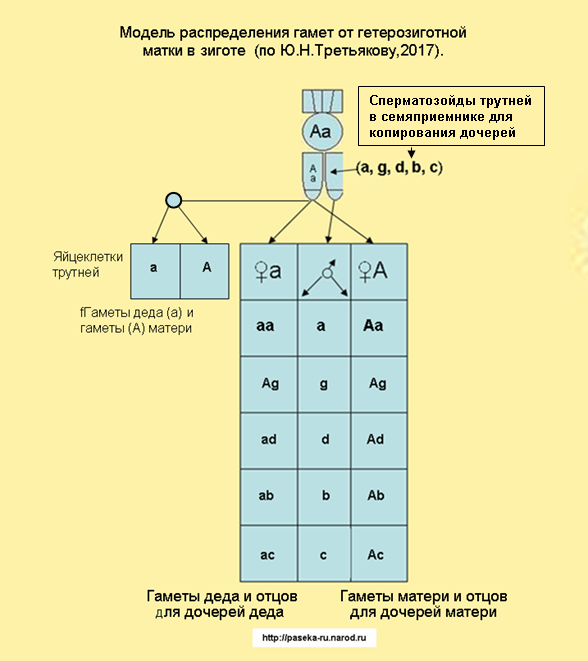
Трутень, как правило, появляется в результате партеногенеза и имеет только один набор хромосом. Он не имеет одинаковых или разных пар аллелей, его называют гемизиготной особью, поскольку свое развитие получает от материнской неоплодотворенной яйцеклетки, Однако при близкородственном спаривании матка может получить от трутня или трутней сперматозоиды одинаковых рецессивных аллелей. Тогда такая матка может откладывать диплоидные яйца, то есть, оплодотворенные, из которых могли бы развиваться трутни с двойным набором хромосом.
При нормальной откладке маткой яиц, половые аллели сперматозоидов трутня или трутней комбинируются с яйцеклетками матки, образуя различные сочетания гамет в потомстве дочерей. Для расчета сочетаний различных типов гамет английский генетик Р. Пеннет предложил использовать решетку, в которой по горизонтали записываются отцовские гаметы, а по вертикали - материнские, в образующихся квадратах - сочетания гамет, т.е. в оплодотворенных яйцеклетках будут женские особи - матки или рабочие пчёлы. Например, в решетке (рис. 8) по вертикали - А,в - яйцеклетки матки пчел, а по горизонтали - а, б, В, г, с - половые клетки разных трутней, в квадратах сочетание материнских и отцовских гамет в зиготах для рабочих пчёл или для молодых маток.
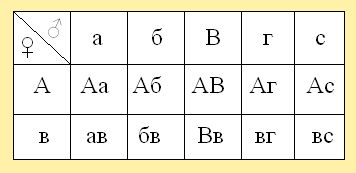
Рис. 8. Решетка по Р. Пеннету с сочетаниями материнских ( Ав) и отцовских ( Ав) и отцовских ( а, б, В, г, с) гамет, где в квадратах - условные женские особи. а, б, В, г, с) гамет, где в квадратах - условные женские особи.
Используя данные из решетки Р. Пеннета, можно представить эти комбинации в моделях яиц и в моделях особей пчел, для первого дочернего поколения F1 - A и по F1 - в (рис. 9).
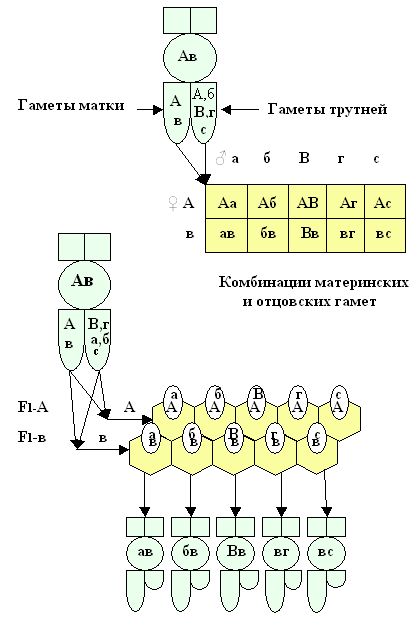
Рис. 9. Модели особей рабочих пчёл из яиц матки (Ав) оплодотворенных гаметами трутней - а,б,В,г,с.
Если нижнюю часть решетки Пеннета раздвинуть в одну или разные стороны, то можно построить в моделях состав особей гнезда пчёл (рис. 10). Как видно из рисунка, что трутни имеют один тип аллеля, а женские особи 2 типа аллелей. Трутней с набором двух одинаковых типов аллелей пчёлы к развитию не допускают, уничтожая в фазе яйца или младшей личинки.
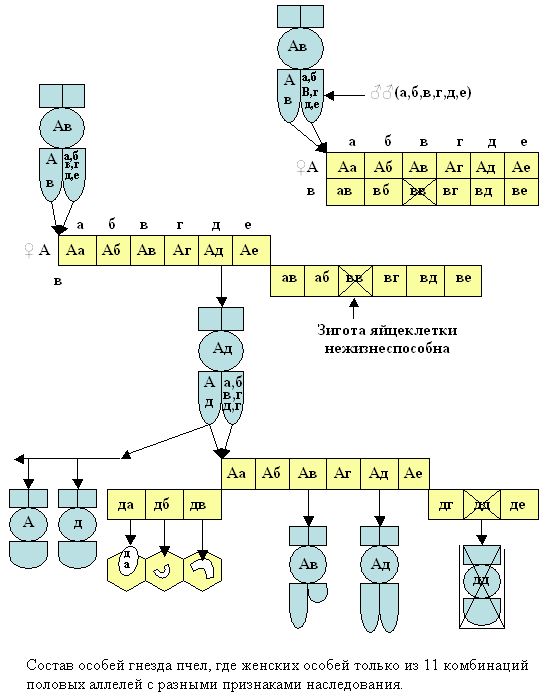
Рис. 10. Варианты построения зигот и моделей особей из решетки Р. Пеннета с одностороннем и разностороннем её частей смещением.
ххххххххх
Законы наследования признаков
Размножение высокопродуктивных семей пчёл можно проводить только при знании закономерностей наследования признаков, которые имеют главное значение в племенной работе. Без знания закономерностей наследования признаков у особей семьи пчел, невозможно выбрать породных особей для скрещивания. Известно, что от чистопородных среднерусских пчёл всегда размножаются среднерусские, от итальянских - итальянские, от серых кавказских - серые горные кавказские и т.д.
Наследственность пчёл проявляется в передаче новым поколениям не только видовых признаков, но также в потомстве наследуются внутривидовые, индивидуальные признаки родителей.
Выбор, тех или иных признаков, при подборе пар для скрещивания проводится путем гибридологического анализа. Сущность метода гибридологического анализа сводится к выбору особей с достаточно четко различимыми признаками, стойко передающихся по наследству (т.е. особо чистопородных). Однако, в племенной работе при выявлении наследственности, селекционеру приходиться встречаться с изменчивостью признаков в потомстве. Похожие внешне предки на родителей, всегда отличаются индивидуальными особенностями, например, характером поведения, медопродуктивностью, зимостойкостью, яйценоскостью и т.д.
Основные законы наследования признаков были впервые разработаны Г. Менделем и опубликованы в 1865г., но до 1900г. оставались, мало известны. С 1900 года они стали широко применяться в растениеводстве и животноводстве.
Известно, что до открытия законов Г. Менделя и до открытия полиандрии пчелиной матки, силезский патер пчеловод Ян Дзержон в 1845 году опубликовал свои наблюдения над окраской тела пчёл при скрещивании в естественных условиях желтых итальянских с темными среднеевропейскими пчелами и открыл развитие трутней из неоплодотворенных яиц.
В пчеловодстве законы наследования стали пригодны к практике с 1925 года, т.е. когда американец Ватсон сделал пригодный шприц для впрыскивания молодой половозрелой матке, определенный объем спермиев трутня, а также была разработана методика инструментального осеменения маток пчел.
Началом изучения законов наследования признаков пчел, в России послужили работы А.С. Михайлова в 1929 – 1930 годах (Комаров, 1935).
Именно искусственное (инструментальное) осеменение маток позволило начать изучение передачи наследственных признаков у пчел. Для изучения наследования стали использовать родительских особей, у которых четко выделился признак наследования с желтой и темной (черной, серой) окраской тела и окраской глаз - белой и черной.
В настоящее время в селекции пчёл выделяют большой объем таких признаков как морфологические, физиологические, биологические, фенотипические.
Скрещивание родительских пар, отличающихся друг от друга одним признаком, называют моногибридным, двумя - дигибридным, тремя - тригибридным, многими - полигибридным.
1. Моногибридное скрещивание
Это такое скрещивание, при котором изучают наследование одного какого-либо признака.
Известно, что схемы наследования отдельных генов пчелы отличаются от обычной схемы менделевского наследования генов и признаков из-за гаплоидности трутней, а также из-за полиандрии маток. И как отмечает Д. Шаскольский (1990), эти схемы похоже на наследование генов «Х»-хромосомы других животных, где у одного пола имеется пара половых «Х»-хромосом, а у другого пола только одна «Х», в то время как у пчелы наследуются гены всех 16 пар хромосом.
Первый закон наследования.
При моногибридном скрещивании у потомства пчёл в первом поколении, от скрещенных родителей матки с трутнем отличающихся друг от друга одним признаком, то потомство женских особей по этим признакам будет единообразно. Такой признак первого поколения называют доминирующим. Это поколение фенотипически единообразно. Например, если для скрещивания возьмем молодую матку желтой окраски и спарим с трутнем черной окраски. Желтая окраска будет доминирующей, обозначим её заглавной буквой А или В, С и т. д., Матка – АА, её желтая окраска является преобладающей. Окраску трутня обозначим малой буквой – а. Построим схему наследования из моделей особей с буквенными обозначениями желтой и черной окраски при скрещивании двух разных по окраске пород пчёл (рис.21.1,2). В результате, после оплодотворения яйцеклеток, зигота получила от матери и отца разные гены А и а. Зигота – (от греческого зиготес - соединенный вместе) – новая яйцеклетка после слияния женской и мужской половых клеток, становится способной к развитию. Такое оплодотворение дает новому организму двойственную наследственность по матери и отцу (трутню) обусловливая жизненность и возможности его приспособиться к новым меняющимся условиям среды обитания.
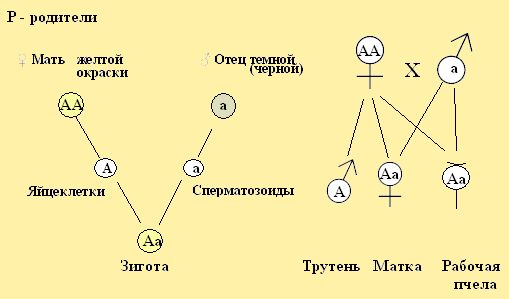
Рис. 21.1. Варианты скрещивания родителей медоносных пчёл (принятые в селекции пчел)
Поскольку в первом поколении желтая окраска у женских особей сохранилась, а черная не проявилась то такой признак называют рецессивным, т.е. черная окраска находится в скрытом виде, и она как признак обозначается малой буквой – а или – в, с и т.д.
Особей получивших от родителей одинаковые аллели гена называют гомозиготными, а разные – гетерозиготными. Следовательно, пчелы, получившие
гены АА; ВВ; СС или аа, вв, сс – будут гомозиготными, а потомство - Аа , Вв, Сс полученных в результате скрещивания – гетерозиготными.
Гены темной (черной) и желтой окраски составляют одну пару противоположных признаков, которые называют аллеломорфными (название устарело) т. е. аллелями.
По окраске, в первом F1 поколении, гетерозиготные особи не отличаются от гомозиготных, т. е. они не отличаются по своему признаку от чистокровных родителей с доминирующей окраской. Трутни же в первом поколении несут признаки присущие матке пчел, как на рис. 21.2.
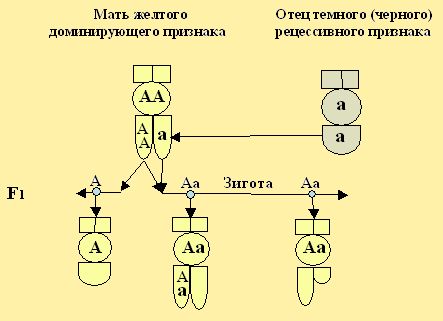
Рис. 21.2. Варианты скрещивания моделей особей медоносных пчёл.
От гомозиготных маток нарождаются трутни одного аллеля гена. Например, матка АА даст трутней А, матка аа даст трутней только а.
Гетерозиготные матки – Аа или Вв во втором F2 дадут трутней – А и а. В первом поколении при скрещивании желтой матки с черным трутнем, мы получим гетерозиготных женских особей с брюшными кольцами доминирующей желтой окраски, т. е. будет наблюдаться единообразие особей. Единообразие F1 поколения сохраниться, если мы спарим матку АА с трутнями, которые могут иметь по одному разному признаку А – доминантному и а – рецессивному (рис. 22). Рецессивный признак, передается по наследству, но подавляется, как уже ранее отмечалось находиться в скрытом виде, не проявляясь у гетерозиготных потомков, полученных при скрещивании.
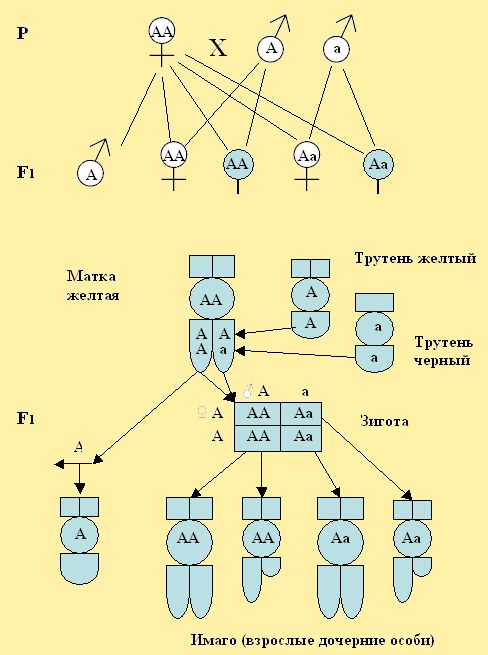
Рис. 22. При скрещивании желтой матки с двумя трутнями имеющих по одному признаку желтой и черной окраски, все особи в F1 единообразны желтые
2. Закон расщепления во втором поколении
При дальнейшем скрещивании между собой гибридов маток и трутней первого поколения F1, которое было единообразно, мы получим расщепление в их потомстве, где во F2 поколении снова появляются женские особи с рецессивными признаками. Такое скрещивание называют анализирующим или возвратным. Для примера возьмем гибридную матку – дочь Аа первого поколения с доминирующей желтой окраской – А и скрытой черной рецессивной окраской – а, и проведем скрещивание с трутнем черной окраски – а, а затем проследим расщепление во втором поколении (рис. 23). При таком варианте скрещивания у пчёл протекает строго определенным числом как женских, так и мужских особей в соотношении 50% желтой окраски и 50% тех и других с черной окраской, что связано с составом родителей по их генам. Например, если плодная матка Аа – желтой окраски была скрещена с трутнем а – черной окраски и отложила 1200 яиц, то из них народятся 600 особей желтой окраски и 600 – черной. Следовательно, от такой матки можно получить рабочих пчел, молодых маток и трутней желтой и черной окраски.
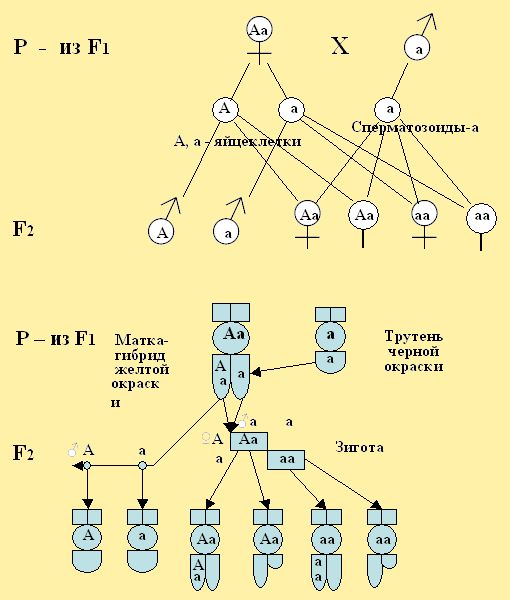
Рис. 23. При скрещивании гибридной матки желтой окраски из первого поколения с трутнем черной (темной, серой) окраски, во втором поколении, можно наблюдать расщепление на особей 50% желтой и 50% черной окраски. (Анализирующее скрещивание).
При анализе наследования признаков у женских гибридных особей гены их могут находиться в скрытом виде (рецессивном), а у трутней скрытых генов нет, т.е. у них проявляются все гены. Следовательно, трутень появляется на свет всегда «чистокровным» так как он развивается из неоплодотворенного яйца. Поэтому, если матка несет трутневые яйца с генами только одного признака, то такая матка тоже «чистокровная», а если от нее будут трутни, к примеру, желтого и черного – то она метис (гибридная). Матка – дочь из первого поколения отличается от своей матери тем, что она развивалась из яйцеклетки с аллелем гена желтого признака, но оплодотворенной сперматозоидом с аллелем гена черного признака. Следовательно, у такой неплодной матки при редукционном делении будут развиваться два сорта яиц, где один сорт яиц несет хромосому, имеющую ген желтой окраски, а другой сорт – хромосома несет ген черной окраски.
При оплодотворении матки – дочери с трутнем черной окраски или желтой окраски, то в её половых органах будут находиться три сорта половых клеток (два сорта яиц и один сорт сперматозоидов).
При моногибридном скрещивании родителей пчёл с доминантными и рецессивными признаками. Например, матку Аа скрестить с трутнями А и а, то потомство будет иметь строго определенное соотношение 3:1 (рис. 24), где 3/4 женских особей будут иметь доминантный признак (желтую окраску) и 1/4 – будут рецессивными (черной окраски). Общее соотношение будет АА : 2Аа : аа, где 1АА : 2Аа – желтой окраски, а 1аа – черной. Формула АА, Аа, аа отражает генотип особей, а – АА, Аа – фенотип. Вероятность, соответствующих комбинаций, можно определить путем алгебраического умножения: (1/2А + 1/2а) х (1/2А + 1/2а) = (1/4АА + 1/4 Аа + 1/4аА + 1/4аа) (яйцеклетки) х (сперматозоиды) = (сочетание гамет при оплодотворении).
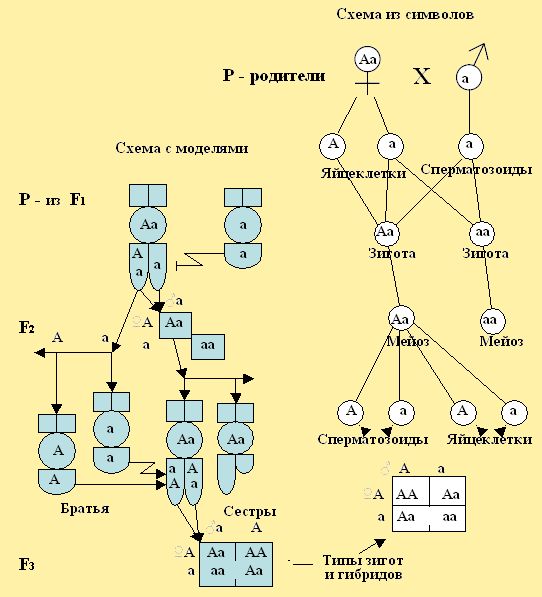
Рис. 24. При скрещивании сестры спермой братьев, где родители несут доминантные и рецессивные признаки, мы получим потомство в строго определенном соотношении: 3/4 особей будут иметь внешние доминантные признаки и 1/4 - рецессивные.
3. Цитологические основы закономерностей наследования
Связь между поколениями особей пчёл при половом размножении осуществляется через половые клетки (гаметы), которые несут материальные наследственные факторы - гены. Гены определяют развитие того или иного признака они расположены в хромосомах. В клетках женских особей пчел, как уже отмечалось ранее, число хромосом составляют 32 (две пары гамет), а у мужских (трутней) 16 (одна пара гамет). Половые клетки женских и мужских особей составляют одну пару хромосом. Две пары хромосом возможны при слиянии женской яйцеклетки и мужской клетки (сперматозоида) в зиготе. Зигота может произойти от слияния женской яйцеклетки и сперматозоида, содержащие одинаковые гены и разные. Например, когда яйцеклетка с геном – А соединится сперматозоидом имеющим ген – А, зигота будет АА, а если яйцеклетка А соединится со сперматозоидом – а то зигота будет Аа. Гомозиготная пчелиная матка АА или аа имеет яйца из гамет А или а, и трутни народившиеся от таких маток будут иметь гаметы А или а.
При рассмотрении цитологических основ моногибридного скрещивания в первом поколении F1 мы наблюдаем в зиготе соединение гамет и расхождение их в мейозе. Это специфическое деление клеток, при котором образуются половые клетки для второго поколения F2 (рис. 25.1). На представленном рисунке 25.2. на моделях пчёл и символах, можно рассмотреть, что у родителей первого поколения F1
АА – желтой окраски и трутня
а – черной окраски все потомство будет единообразно, т.е. – желтое. Во втором поколении F2 произошло расщепление в желтых особях на желтых и черных в соотношении 3:1, где три будут нести желтую окраску, а одна особь – черную.
В генетике гетерозиготная особь носит название гибрид. Потомка, полученного от скрещивания родителей разных пород, называют метисом.
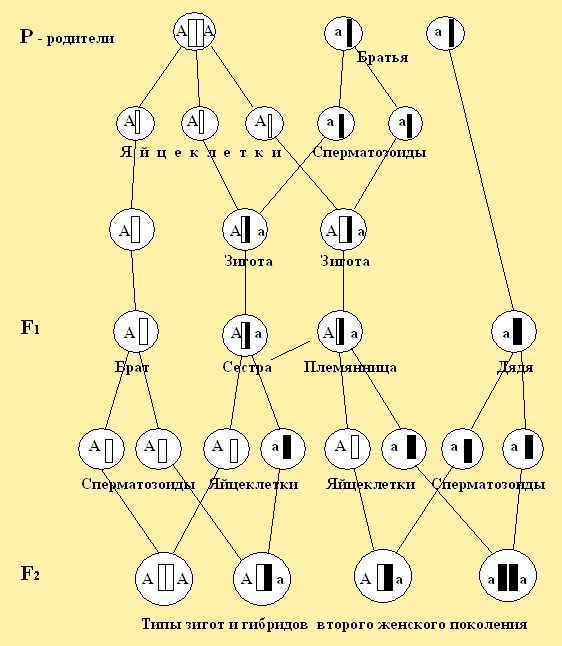
Рис. 25.1. Цитологические основы моногибридного расщепления, где расхождение пар гамет в мейозе и объединение их при оплодотворении в типах зигот потомства пчел.
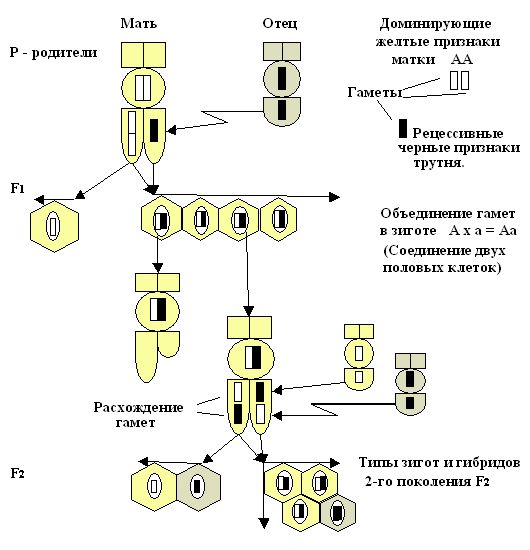
Рис. 25.2. Цитологические основы моногибридного расщепления гамет в потомстве от гомозиготной матки желтого доминирующего признака и гомозиготного трутня черного рецессивного признака (на моделях пчел). Расхождение пар гамет в яйцеклетках при мейозе и объединение их при оплодотворении в типах зигот во втором поколении гибридов.
4.6.1. Дигибридное скрещивание
Второй закон Менделя
Если моногибридное скрещивание позволяет проследить наследование одного признака, что имеет важное, значение в селекции, особенно для начинающих селекционеров (генетиков), то при дигибридном скрещивании можно проследить, как протекает наследование у потомства, имеющие родителей отличающихся друг от друга двумя признаками. Для примера, рассмотрим простые формы скрещивания родителей, где матка пчёл будет иметь желтую окраску и черные глаза, а трутни черной окраски с белыми глазами. Обозначим матку ААВВ, а трутня ав. Желтая окраска тела и черные глаза будут доминирующей окраски, а черная окраска тела и белые глаза будут иметь рецессивные признаки.
При дигибридном скрещивании зигота будет иметь обозначение из четырех букв, из которых две буквы относятся к яйцеклеткам и две - к сперматозоидам. В этом скрещивании первый закон наследования сохраняет свою силу, т.к. все женские особи, будут в первом поколении единообразны, так как у них проявляются доминирующие признаки, а у трутней выявляются признаки присущие матке пчел.
В первом поколении все рабочие пчёлы или матки будут желтые, черноглазые (АаВв), а трутни черноглазые, желтые (АВ) от матки ААВВ и трутней ав.
Расщепление будет наблюдаться во втором поколении, когда матка АаВв после скрещивания, например с трутнем ав станет откладывать четыре типа яиц (по цвету они неотличимы), из которых можно получить четыре типа женских особей и четыре типа мужских (трутней) в соотношении 25% каждого типа из четырех (рис.26). В цифровых соотношениях - 1:1:1:1.
Если взять одну матку или несколько маток из первого поколения и оплодотворить с трутнями из второго имеющих типы гамет АВ, Ав, аВ, ав, то в этом случае варианты скрещивания могут быть различны. От такого скрещивания родителей во втором поколении мы получим более сложную комбинацию из 16 типов зигот женских особей с формулой 9АВ:3Ав:3аВ:1ав. Алгебраически, это квадрат двучлена (3+1)2 = 32 + 2 x 3 + 12. В этом случае закон Менделя гласит: расщепление по каждой паре признаков идет независимо от других пар признаков.
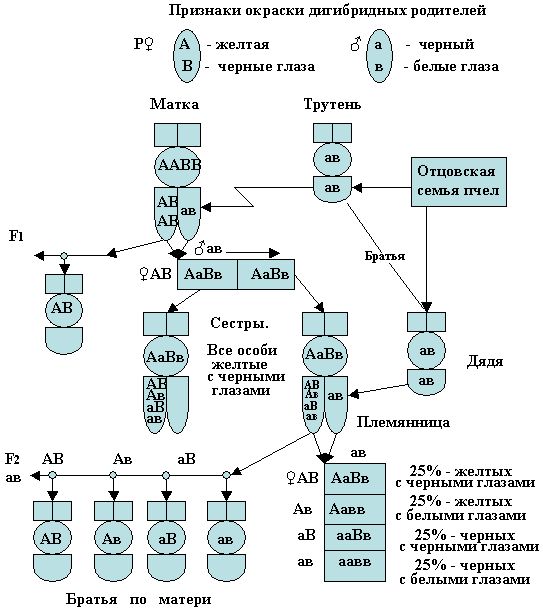
Рис. 26. При дигибридном скрещивании гомозиготных родителей пчел, мы получим гетерозиготных особей по обоим доминирующим признакам, а во втором F2 поколении можно наблюдать расщепление признаков на 4 типа зигот в
Третий закон Менделя
Признаки, полученные от одного из родителей, не связаны между собой, не передаются всегда вместе, могут разъединяться и каждый из них свободно комбинируется с признаками, полученными от другого родителя.
Согласно третьему закону наследования – это закон независимости комбинаций генов расположенных в разных парах хромосом. Исходя из законов Менделя в основе дигибридного скрещивания, всегда будет находиться моногибридное расщепление в соотношении 3:1 (при наличии доминирования), для дигибридов это будет (3:1)2, для тригибридов – (3:1)3, для n–й степени гибридности – (3:1)n.
Цитологические основы дигибридного скрещивания родителей пчел
При рассмотрении цитологических основ дигибридного скрещивания в первом поколении (F1) мы можем наблюдать соединение гамет в зиготе и расхождение их в мейозе. Процессы независимого распределения генов, наблюдается при расхождении их во втором поколении, которые совершаются в половых клетках при их созревании и при оплодотворении.
В диплоидном наборе хромосом расположены аллельные гены. Отдельные хромосомы могут свободно комбинироваться одна с другой при образовании гамет, а в половых клетках матки АаВв не могут быть вместе, например, А и а или В и в, но гены могут комбинироваться одна от другой при условии, когда гены парного признака находятся в хромосомах различных пар.
Так, например, у матки пчёл ААВВ (желтой окраски с черными глазами) гены АА можно представить в виде палочкообразных ( ) - желтой окраски, а гены ВВ – в сферическом виде ( ) - желтой окраски, а гены ВВ – в сферическом виде ( ) – черной окраски. У самцов ав – трутни (черной окраски с белыми глазами) - гены а будут иметь палочкообразный вид ( ) – черной окраски. У самцов ав – трутни (черной окраски с белыми глазами) - гены а будут иметь палочкообразный вид ( ) – черной окраски, а гены в - сферическую форму ( ) – черной окраски, а гены в - сферическую форму ( ) – белой окраски (рис.27). В результате мейоза в половых органах матки (яичниках) будут нарождаться яйцеклетки с одной парой генов АВ – (). При оплодотворении матки с яйцеклетками АВ () трутнем ав () в гетерозиготе по двум признакам АаВв в каждой паре хромосом будут находиться разные гены одной пары аллелей. ) – белой окраски (рис.27). В результате мейоза в половых органах матки (яичниках) будут нарождаться яйцеклетки с одной парой генов АВ – (). При оплодотворении матки с яйцеклетками АВ () трутнем ав () в гетерозиготе по двум признакам АаВв в каждой паре хромосом будут находиться разные гены одной пары аллелей.
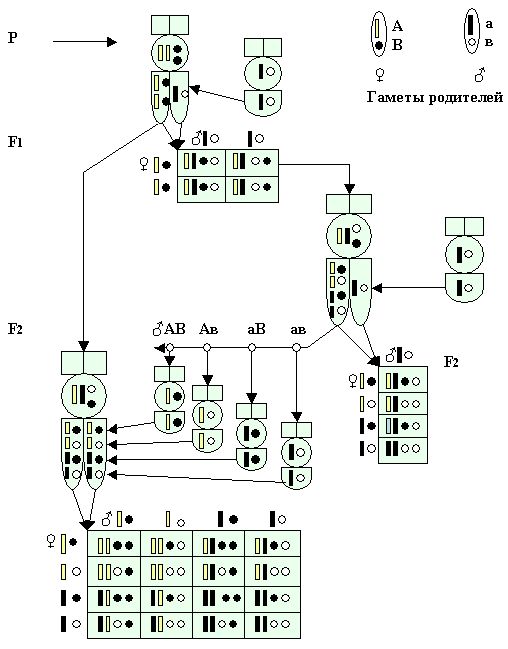
Рис. 27. Цитологические основы дигибридного скрещивания с распределением гамет пчёл в потомстве и типах гибридов зигот первого F1 и второго F2 поколения.
При мейозе у гибридов первого поколения F1 в разном количестве образуются четыре сорта гамет. Если, например, к одному полюсу отходит черная палочковидная хромосома, то из другой пары с одинаковой долей вероятности может отойти желтая палочка и т.д. В результате оплодотворения матки АаВв с трутнем ав и развития второго поколения гибридов F2 будет одинаковое численное образование четырех категорий зигот женских особей. А также из неоплодотворенных яиц могут народиться четыре категорий трутней – носителей сперматозоидов этих категорий.
Если все четыре категории парных генов трутней при случайных половых встречах или при искусственном осеменении попадут в семяприемник гетерозиготной молодой матки АаВв первого поколения имеющей четыре категории парных гамет в яйцеклетках, то в результате расхождения генов в мейозе и слиянии, по двум парам признаков, в зиготе, образуются женские особи различающиеся по двум парам признаков в 16 различных комбинациях (рис. 28).
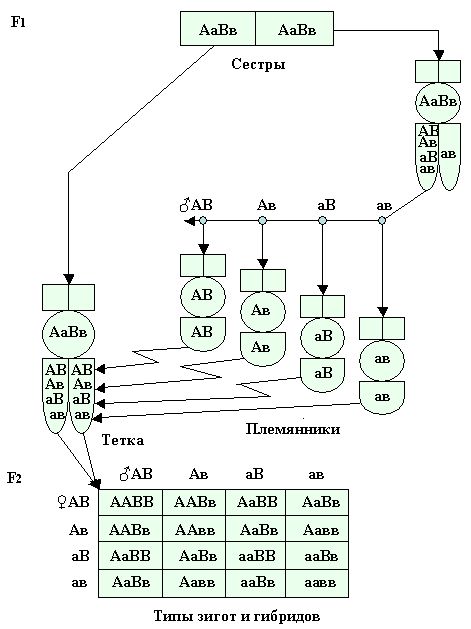
Рис. 28. При дигибридном скрещивании гетерозиготных родителей, пчелиной матки из F1 с трутнями от сестры из F1, мы получим в F2 16 различных комбинаций типов зигот гибридов, а из них 16 различных комбинаций сводных сестер.
4.6.2. Тригибридное скрещивание родителей пчел
При дигибридном скрещивании родители (матка и трутень) отличаются друг от друга двумя различными признаками аллельных ген. Однако, в селекции пчёл в большинстве случаев необходимо проследить, как будет передаваться наследственность от родителей потомкам, если родители отличаются тремя и более наследственными признаками.
Известно, что каждая новая женская особь семьи пчёл возникает от слияния двух половых клеток (яйца и сперматозоида), в результате половину своих генов получает от матери, а другую половину генов от трутня (отца). От такого слияния образуются парные гены. Парные гены называют аллельными, а гены – это реально существующие, независимо комбинирующиеся и расщепляющиеся единицы наследственности.
Для составления всех типов половых клеток любого скрещивания (от дигибридного, тригибридного и т. д.) необходимо последовательно разъединять каждую пару аллельных генов родителей и сочетать их с ранее отщепившимися. Например, возьмем разные пары контрастных альтернативных признаков из моногибридного, дигибридного, тригибридного и полигибридного - черыре пары признаков и проследим, как разъединяются и соединяются парные гены, для образования половых клеток (рис. 29).
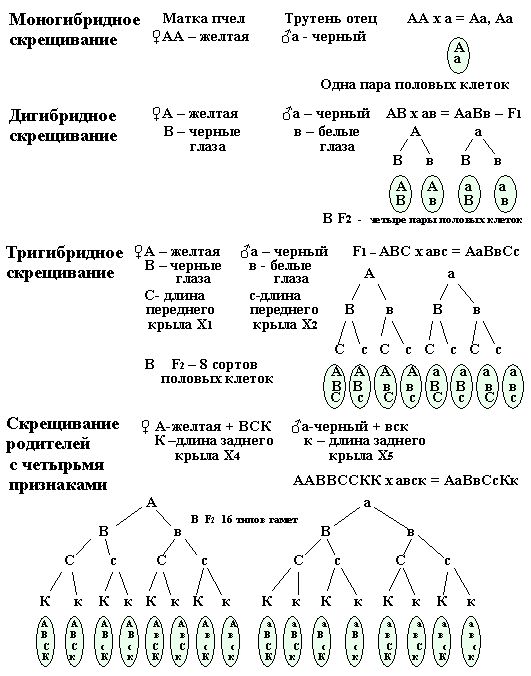
Рис. 29. Разъединение и соединение контрастных признаков парных аллельных ген при дигибридном, тригибридном и с четырьмя парами – при скрещивании родителей пчел.
При тригибридном скрещивании с тремя парами признаков, первое поколение однородно и имеет доминирующие признаки родителей. В первом поколении с одним трутнем можно получить 4 комбинации, с 4–мя – 16 комбинаций, а во втором поколении наблюдается более сложных характер, так как образуется 8 сортов гамет, которые при оплодотворении дают 64 комбинации (рис. 30) из 8 фенотипов и 27 генотипов. Расщепление по фенотипу во втором поколении проходит в отношении:
27АВС : 9АВс : 9АвС : 9аВС : 3Авс : 3аВс : 3авС : 1авс
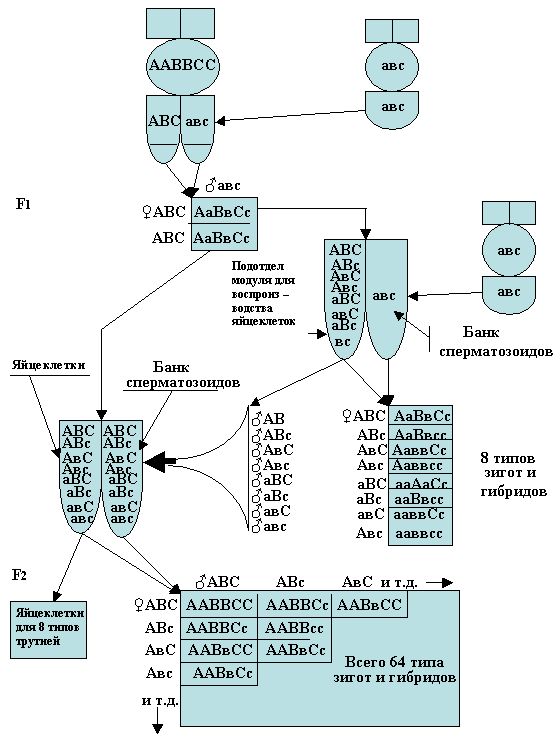
Рис. 30. Тригибридное скрещивание родителей пчёл с тремя парами контрастных аллельных признаков в первом F1 и втором F2 поколении.
4.6.3. Полигибридное скрещивание
Полигибридное скрещивание маток и трутней пчёл отличается от тригибридного скрещивания тем, что эти родители имеют не три признака отличия, а гораздо больше, т.е. родительские особи пчёл при скрещивании различаются друг от друга большим числом пар признаков и сортов гамет во втором поколении.
При полигибридном скрещивании расщепление проходит сложнее и сильнее возрастает комбинативная особенность, ибо при комбинировании генов каждый член аллельной пары может комбинироваться с любым числом остальных пар.
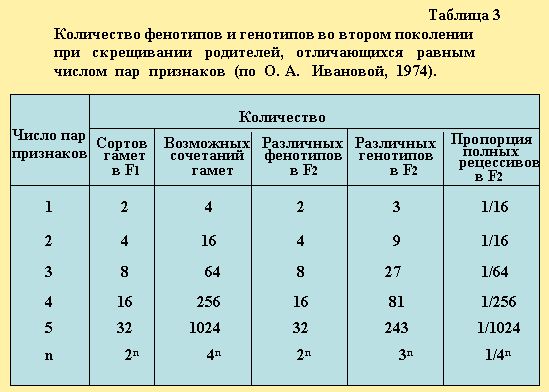
Для наглядности воспользуемся таблицей 3 (Иванова.1974) в которой приведены данные сортов гамет от F1 до Fn с указанием количества фенотипов и генотипов, в том числе генотипов второго поколения при скрещивании родителей отличающихся равным числом пар, различных признаков. Например, если принять n = 6, тогда число пар признаков равное 6 сортов гамет первого поколения (F1) будет составлять 2 = 64, а возможность сочетания гамет 46 = 4096, в этом случае число различных фенотипов второго поколения F2, будет 26 = 64, а различных генотипов в F2 - 36 = 729.
Используя таблицу 3 (по О.А. Ивановой,1974) можно решать различные варианты скрещивания с любым числом аллельных пар признаков.
В селекционной работе пчеловодства используют целый ряд различных генетических свойств и качество пчёл по количественным и мерным признакам. Пчелы, как объект племенной работы в значительной степени отличаются от других сельскохозяйственных животных, что усложняет селекцию. Отличие состоит в том, что пчелиная матка (как уже отмечалось выше) в естественных условиях спаривается с трутнями в воздухе при полете, а каждый трутень после акта спаривания мгновенно погибает, а в дополнение к этому в спаривании участвуют несколько трутней, количество которых может составлять от 4 до 10 и даже до 15 (о 15 трутнях упоминает Н.И. Кривцов, 1995). В этом количестве трутней, каждый из них может нести свой набор отличительных признаков, а потому в селекции пчёл используется инструментальный метод искусственного оплодотворения, которое контролируется по более четким отличительным признакам родительских пар. Однако, и в этом методе, используют не одного трутня, а несколько от 4 до 6, при одном впрыскиваний, а количество таких впрыскиваний спермы необходимо от 2 до 3 раз.
Следовательно, и в этом случае, могут быть отклонения от теоретических, чистых спариваний с трутнями из одной семьи.
Расчет возможных теоретических комбинаций гамет у пчёл можно определить заранее. Если учитывать, что у пчелиной матки в половых органах по одной паре генов в каждой из 16 пар хромосом, возможно появление 2 в 16 степени разных типов гамет и 4 в 16 степени возможных вариантов их сочетаний, что дает неограниченные возможности наследственной изменчивости.
Развитие признаков организма определяется взаимодействием множества генов, а каждый ген обладает множественным действием. Множественное действие генов оказывает влияние не на один, а на ряд признаков организма. Генотип организма связан с хромосомным аппаратом, т.е. с ДНК, а ДНК каждого гена, является передатчиком генетической информации, которая закодирована и «переписывается» (транскрибируется) с генной ДНК на молекулу информационной РНК. В свою очередь РНК синтезируется в ядре и переходит в цитоплазму к рибосомам эндоплазматической сети
. |
